渤海灣太平洋牡蠣殼體記錄的冬季停止分泌/春季恢復(fù)生長(zhǎng)的最低水溫
2012-09-20 00:46:30范昌福裴艷東王海峰
地球?qū)W報(bào) 2012年6期
關(guān)鍵詞:生長(zhǎng)
范昌福, 王 宏, 裴艷東, 王海峰
1)中國(guó)地質(zhì)科學(xué)院礦產(chǎn)資源研究所, 北京 100037;2)國(guó)土資源部成礦作用與資源評(píng)價(jià)重點(diǎn)實(shí)驗(yàn)室, 北京 100037;3)中國(guó)地質(zhì)調(diào)查局天津地調(diào)中心, 天津 300170
渤海灣太平洋牡蠣殼體記錄的冬季停止分泌/春季恢復(fù)生長(zhǎng)的最低水溫
范昌福1,2), 王 宏3), 裴艷東3), 王海峰3)
1)中國(guó)地質(zhì)科學(xué)院礦產(chǎn)資源研究所, 北京 100037;2)國(guó)土資源部成礦作用與資源評(píng)價(jià)重點(diǎn)實(shí)驗(yàn)室, 北京 100037;3)中國(guó)地質(zhì)調(diào)查局天津地調(diào)中心, 天津 300170
河口海岸地區(qū)廣泛生長(zhǎng)的牡蠣殼體, 作為氣候環(huán)境變化的一種載體, 是研究河口淺海環(huán)境演化和海陸相互作用的理想對(duì)象。本文以渤海灣地區(qū)廣泛發(fā)育的太平洋牡蠣殼體為研究對(duì)象, 根據(jù)現(xiàn)生活牡蠣殼體記錄的氧同位素變化幅度和生長(zhǎng)水體的溫度、鹽度, 定量地計(jì)算了該地區(qū)Crassostrea gigas在冬季停止分泌/春季恢復(fù)生長(zhǎng)的最低水溫為5.5℃, 生長(zhǎng)季節(jié)為3月至12月。
生物殼體; 停止分泌; 氧同位素; 水溫; 渤海灣
高分辨率氣候和環(huán)境變化研究對(duì)了解過(guò)去環(huán)境變化中突發(fā)事件和未來(lái)環(huán)境變化的預(yù)測(cè)具有重要意義。可信的氣候預(yù)測(cè)模型的建立, 需要來(lái)自不同地區(qū)環(huán)境類型下的高精度、長(zhǎng)周期氣候記錄的互相校驗(yàn)(Sch?ne et al., 2005), 包括陸地、海洋和陸海交互地帶的海岸河口地區(qū)的各種氣候記錄。
近幾十年以來(lái), 我國(guó)全新世高分辨率古氣候環(huán)境重建研究取得了大量的重要成果, 但絕大部分成果集注于內(nèi)陸和深海地區(qū), 因而不能代表同時(shí)期陸海交互地帶的氣候信息(Sch?ne et al., 2004, 2005)。作為連接內(nèi)陸與海洋的海岸帶地區(qū), 由于復(fù)雜的相互作用和頻繁的地質(zhì)環(huán)境變化, 難以保存連續(xù)的長(zhǎng)時(shí)間序列地質(zhì)環(huán)境記錄, 因此很難獲得可與內(nèi)陸及深海地區(qū)進(jìn)行對(duì)比的氣候環(huán)境記錄。
牡蠣是河口及淺海區(qū)一種常見(jiàn)的半咸水雙殼類軟體生物, 屬?gòu)V溫、廣鹽性, 從低緯度至中高緯度的河口淺海環(huán)境都適合其生長(zhǎng); 牡蠣從白堊紀(jì)一直繁衍存活至今, 其殼體幾乎全部由方解石組成。牡蠣個(gè)體的壽命較長(zhǎng), 很多牡蠣個(gè)體的生長(zhǎng)年齡超過(guò)15 a或更多(張璽等, 1956)。由于牡蠣的時(shí)空分布范圍廣、生長(zhǎng)壽命長(zhǎng)、生長(zhǎng)環(huán)境獨(dú)特等特性決定了其殼體是一種良好的環(huán)境記錄載體, 是研究氣候環(huán)境變化, 特別是河口淺海環(huán)境演化和海陸相互作用的理想對(duì)象(王強(qiáng)等, 1991; 耿秀山等, 1991; Wang et al., 1995; Kirby et al., 1998; 薛春汀, 2003; 張忍順,2004; Surge et al., 2001; Carbotte et al., 2004; 王宏,1996; 王宏等, 2006; Zhang et al., 2007; Brigaud et al.,2008; Wisshak et al., 2009; Ullmann et al., 2010; 李冬玲等, 2009; 商志文等, 2010; 范昌福等, 2005a, b,2006, 2007, 2008, 2010a, b, c, d; Fan et al., 2011; 王海峰等, 2011; 方晶等, 2011)。
太平洋牡蠣(Crassostrea gigas; Thunberg, 1793),生長(zhǎng)于水深5~40 m的淺水區(qū), 濾食水中浮游生物,殼體之間互相膠結(jié)生長(zhǎng)而成礁體。喜好生長(zhǎng)于硬質(zhì)基底, 如巖石、殘骸碎屑和生物殼體等, 但隨著生物的進(jìn)化, 它們也能夠生長(zhǎng)于硬的泥質(zhì)或泥沙質(zhì)基底之上。其廣溫、廣鹽性使其能夠在全世界范圍內(nèi)廣泛繁殖(Cognie, 2006; Diederich, 2005)。它們可生長(zhǎng)的鹽度范圍為 10×10-12~42×10-12、溫度范圍為4℃~35℃, 短時(shí)間內(nèi)甚至能夠忍受–5℃的水溫(Nehring, 2006)。然而, 過(guò)低的水體鹽度將影響牡蠣個(gè)體的配子發(fā)育(gametogenesis)。
渤海灣西岸地區(qū)自早全新世最大海侵以來(lái)(林防等, 2004), 發(fā)育了由C. gigas及它的簇生群體構(gòu)成的規(guī)模巨大、數(shù)量眾多的埋藏牡蠣礁體(圖1), 是該地區(qū)一個(gè)顯著特征和重要地質(zhì)現(xiàn)象。牡蠣礁與渤海灣西岸貝殼堤平原的貝殼堤(王宏等, 2000), 是華北平原東部沿海低平原自中全新世逐漸成陸過(guò)程中的兩類重要產(chǎn)物。它們的形成、規(guī)模、時(shí)間跨度及所蘊(yùn)含的豐富地質(zhì)環(huán)境變化信息, 具有重要的研究?jī)r(jià)值(王宏等, 2006)。
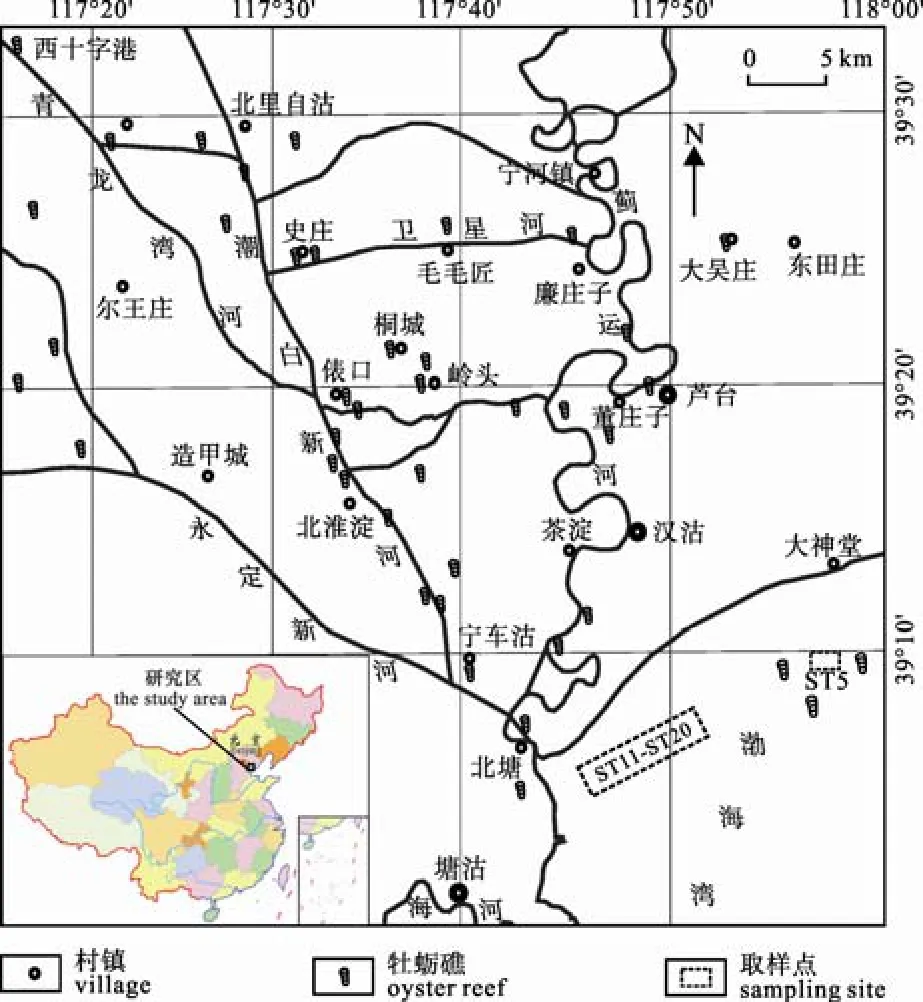
圖1 渤海灣全新世埋藏牡蠣礁體及大神堂海區(qū)活牡蠣礁體分布位置圖(據(jù)王宏等, 2006; 范昌福等, 2010d)Fig. 1 Map showing the distribution of buried and living oyster reefs on the northwest coast of Bohai Bay and the sampling site for this study(after WANG et al., 2006; FAN et al., 2010d)
但是在利用牡蠣殼體進(jìn)行古氣候環(huán)境恢復(fù)重建之前, 需要對(duì)其自身進(jìn)行深入細(xì)致的了解, 以掌握其記錄環(huán)境信息的方式、可靠性和連續(xù)性等。我們?cè)鴮?duì)渤海灣西北岸地區(qū)的太平洋牡蠣殼體的韌帶部位特征生長(zhǎng)層進(jìn)行了詳細(xì)的研究, 共在殼體韌帶槽部位發(fā)現(xiàn)了兩類具有季節(jié)指示意義的特征生長(zhǎng)層(范昌福等, 2010a; Fan et al., 2011), 其中對(duì)應(yīng)于殼體韌帶槽表面凹溝的殼體剖面內(nèi)灰色半透明生長(zhǎng)層,具有一個(gè)年生長(zhǎng)周期內(nèi)最重的氧同位素比值, 是晚秋至早春季節(jié)水溫較低時(shí)期形成的緩慢致密生長(zhǎng)層(范昌福等, 2010a; Fan et al., 2011)。然而, 這一特征生長(zhǎng)層是否指示了其在冬季低溫時(shí)期的生長(zhǎng)間斷?如果C. gigas在冬季低溫季節(jié)停止分泌殼體, 則其停止分泌殼體的最低水溫是多少?這將為使用牡蠣殼體進(jìn)行古氣候環(huán)境高分辨率重建研究提供必要的基礎(chǔ)信息。
1 材料與方法
本次選擇的是2005年5月26日于天津北部海區(qū)活牡蠣礁體中(大神堂, 圖 1)采集的一個(gè)活
C. gigas殼體(ST5), 去除殼體內(nèi)的有機(jī)質(zhì)后, 選擇殼體相對(duì)較大、更適宜于進(jìn)行貝殼年輪研究的左殼殼體(Surge et al., 2001), 沿殼體最大生長(zhǎng)線切割,對(duì)切割后的殼體剖面進(jìn)行打磨和拋光, 最終使殼體切割縱剖面內(nèi)的生長(zhǎng)層清晰可辨(范昌福等, 2010a)。
通過(guò)對(duì)牡蠣殼體韌帶槽表面記錄的因冬季低溫和春季產(chǎn)卵而形成的內(nèi)凹層和外凸層中部的小凹痕,以及殼體切割剖面內(nèi)對(duì)應(yīng)的灰色半透明生長(zhǎng)層的仔細(xì)辨認(rèn)和計(jì)數(shù), 獲得牡蠣殼體的生長(zhǎng)年輪信息(范昌福等, 2010a; Fan et al., 2011)。
在獲取殼體生長(zhǎng)年輪信息的基礎(chǔ)上, 根據(jù)殼體剖面內(nèi)特征生長(zhǎng)層的位置, 使用0.5 mm直徑鉆頭的微型鉆機(jī), 沿殼體生長(zhǎng)方向垂直于殼體剖面鉆取樣品, 獲取約1 mg碳酸鈣粉末, 充分混合均勻后分成三份并分別進(jìn)行測(cè)定, 結(jié)果取三次測(cè)量的平均值。
樣品測(cè)試在德國(guó)漢諾威萊布尼茲應(yīng)用地質(zhì)科學(xué)研究所(GGA)同位素與年代實(shí)驗(yàn)室采用連續(xù)流方法測(cè)定, 運(yùn)用連接Delta XP質(zhì)譜儀的GasBench II裝置自動(dòng)進(jìn)樣, 在 75℃下加入 5滴無(wú)水磷酸, 充分反應(yīng)后測(cè)定樣品的δ18O、δ13C比值, 樣品重復(fù)測(cè)量誤差一般小于0.1‰。儀器的δ18O和δ13C測(cè)試精度分別為0.13‰和0.06‰。
另于 2007年秋季對(duì)活牡蠣礁體生長(zhǎng)區(qū)不同鹽度值的水體進(jìn)行取樣, 使用 YSI85型多參數(shù)水質(zhì)監(jiān)測(cè)儀現(xiàn)場(chǎng)測(cè)定水樣的鹽度值, 用10 mL棕色玻璃瓶現(xiàn)場(chǎng)直接裝滿后即刻密封, 置于陰涼處保存、待測(cè)。水樣δ18O的測(cè)定在南京大學(xué)內(nèi)生金屬礦床成礦機(jī)制研究國(guó)家重點(diǎn)實(shí)驗(yàn)室采用水平衡法完成, 分析儀器為Deltaplus XP。取0.3 mL水樣直接在惰性氣體環(huán)境下與0.2%的二氧化碳?xì)怏w進(jìn)行平衡18 h, 測(cè)試精度小于0.1‰。
2 結(jié)果
2.1 研究區(qū)牡蠣生長(zhǎng)環(huán)境
研究區(qū)位于渤海灣西北部, 海水溫度、鹽度的變化除了取決于氣象因素外, 大陸徑流、陸地?zé)嵝?yīng)以及海灣內(nèi)海水流動(dòng)也是重要的影響因素。春季受太陽(yáng)輻射逐漸加強(qiáng), 水溫和氣溫處于增溫階段,由于近岸海水受陸地?zé)彷椛涞挠绊? 致使其增溫速度較外海快。另外, 春季為江河枯水期, 入海徑流小,加之空氣干燥及風(fēng)的作用, 使海水蒸發(fā)量增大, 鹽度增高, 春季近岸的溫度、鹽度較外海高。夏季溫度普遍比春季升高 12~13℃, 而鹽度升高不大。秋季和前兩個(gè)季節(jié)相反, 秋季為降溫期, 太陽(yáng)輻射逐漸減弱, 氣溫下降, 海水向大氣的回輻射加強(qiáng), 海水處于降溫過(guò)程。特別是近岸淺水區(qū), 由于陸地降溫迅速, 海水向陸地的熱量輸送加強(qiáng), 使近岸海水降溫較快。冬季海水溫度處于最低值, 近岸受陸地迅速降溫影響, 1月份近岸海水溫度達(dá)–5℃, 海水鹽度較高, 沿岸海水鹽度值大于 33×10-12(天津市海岸帶和海涂資源綜合調(diào)查領(lǐng)導(dǎo)小組辦公室, 1987)。
根據(jù)塘沽驗(yàn)潮站 2005—2009年實(shí)測(cè)數(shù)據(jù)計(jì)算的月平均結(jié)果(表 1), 研究區(qū)水溫(SST)呈規(guī)律性變化, 1月份水溫最低, 8月份水溫最高; 水體鹽度(SSS)變化與降雨量(P)有關(guān), 夏季降雨量顯著增強(qiáng), 水體鹽度值明顯降低(圖2)。
2.2 研究區(qū)水體δ18O與鹽度的關(guān)系
根據(jù)研究區(qū)現(xiàn)生活牡蠣礁體生長(zhǎng)水體不同鹽度值的水樣氧同位素比值測(cè)定結(jié)果(表2), 對(duì)其進(jìn)行線性擬合后(Kirby et al., 1998; Surge et al., 2001; Simstich et al., 2005)(圖3), 獲得渤海灣地區(qū)現(xiàn)生水體鹽度-氧同位素組成定量關(guān)系式:
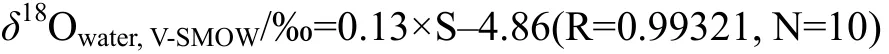
式中δ18Owater,V-SMOW為水體氧同位素組成, S為水體鹽度; 下標(biāo) V-SMOW 表示體積分?jǐn)?shù)以SMOW(標(biāo)準(zhǔn)平均海水)為標(biāo)準(zhǔn), 單位為‰。
由于水樣的氧同位素測(cè)定結(jié)果是以 SMOW 為標(biāo)準(zhǔn), 而殼體碳酸鈣粉末樣品的氧同位素結(jié)果則是以 PDB為標(biāo)準(zhǔn)的, 二者之間存在定量的轉(zhuǎn)換關(guān)系(Rye et al., 1980)。本文采用以下的定量轉(zhuǎn)換關(guān)系式(Gonfiantini et al., 1995):
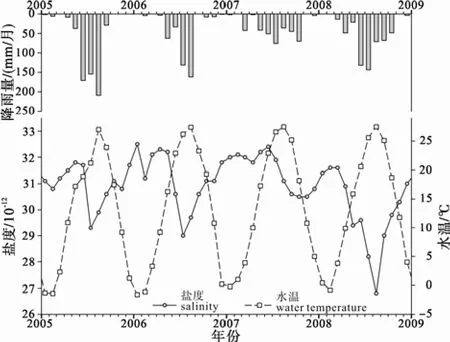
圖2 渤海灣西北岸地區(qū)2005—2009年的月均水溫、鹽度和降雨量(據(jù)Fan et al., 2011)Fig. 2 Average monthly sea surface temperature, sea surface salinity and precipitation during 2005 to 2009 on the northwest coast of Bohai Bay(after Fan et al., 2011)
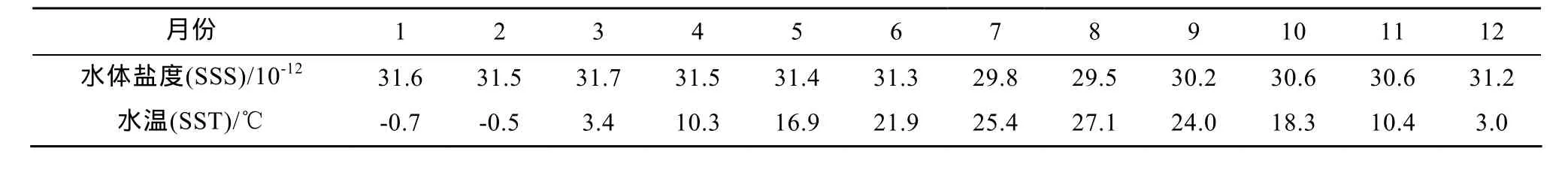
表1 渤海灣西北岸地區(qū)2005–2009年月均水溫與鹽度值Table 1 Average monthly sea surface temperature and sea surface salinity during 2005 to 2009 on the northwest coast of Bohai Bay

表2 渤海灣現(xiàn)生活牡蠣礁體生長(zhǎng)區(qū)不同鹽度的水體δ18OV-SMOWTable 2 Brackish waters and their corresponding δ18OV-SMOW values in the living oyster reef area of Bohai Bay
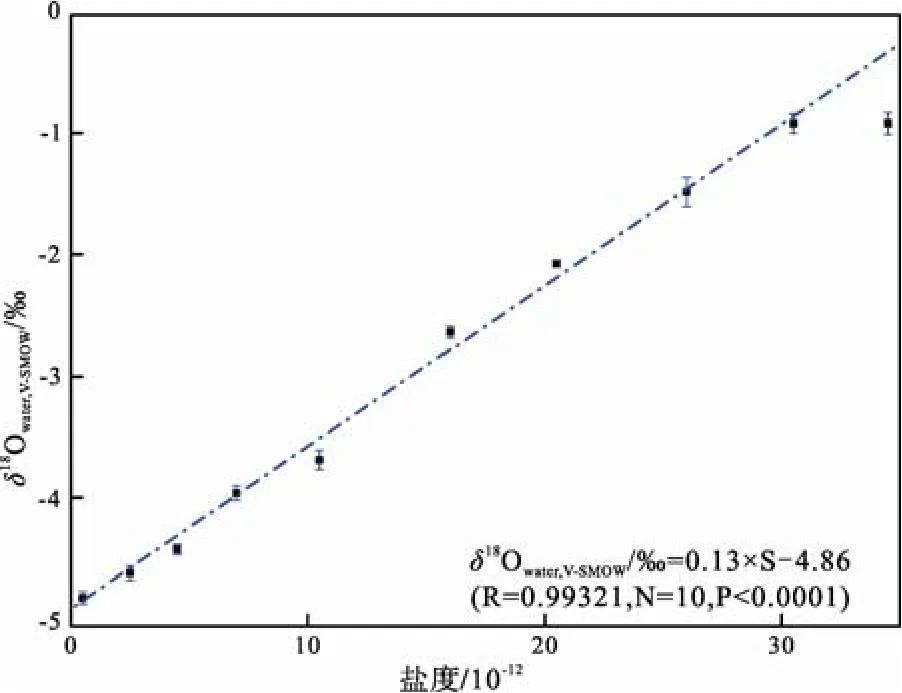
圖3 渤海灣現(xiàn)生活牡蠣礁體生長(zhǎng)區(qū)水體氧同位素組成與鹽度關(guān)系圖Fig. 3 Relationship between δ18OV-SMOW and salinity of the brackish water in the living oyster reef area of Bohai Bay
δ18OV-PDB=(δ18OV-SMOW–30.91)/1.0309將SMOW標(biāo)準(zhǔn)轉(zhuǎn)換為 PDB標(biāo)準(zhǔn)之后, 渤海灣地區(qū)的水體鹽度與水體δ18O的變化關(guān)系可表達(dá)為:δ18Owater,V-PDB/‰=0.12×S–26.7。即水體鹽度值每增加1×10-12, 水體氧同位素比值(PDB標(biāo)準(zhǔn))增加0.12‰。
2.3 活牡蠣殼體記錄的年際δ18O變化幅度
在ST5殼體剖面內(nèi), 共有4個(gè)對(duì)應(yīng)于殼體韌帶槽表面凹槽的灰色半透明生長(zhǎng)層(圖 4中箭頭所示),它們具有一個(gè)年生長(zhǎng)周期內(nèi)最重的δ18O值, 形成于冬季低溫季節(jié)(范昌福等, 2010a; Fan et al., 2011)。在每?jī)蓚€(gè)冬季低溫季節(jié)生長(zhǎng)層之間、具有最輕δ18O值的生長(zhǎng)層, 形成于夏季最高水溫季節(jié)。
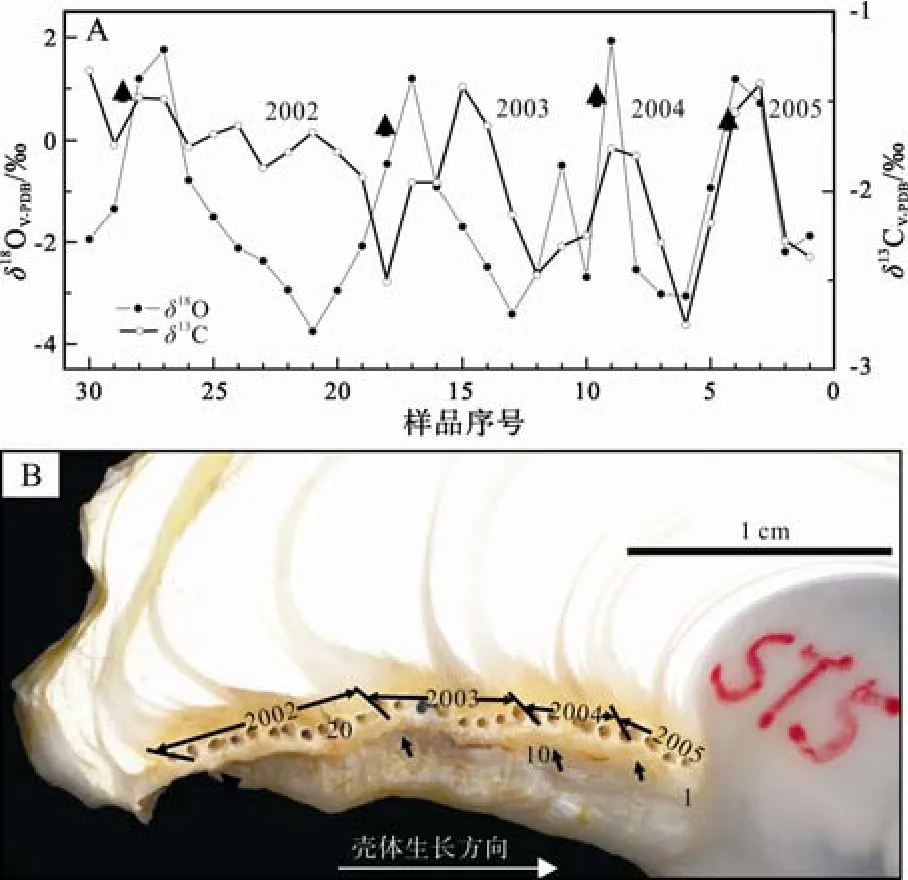
圖4 ST5牡蠣殼體同位素剖面與殼體剖面內(nèi)連續(xù)微層樣品取樣位置Fig. 4 Micro-sampling positions and isotope values of the left valve of the modern living shell ST5箭頭指示冬季低溫季節(jié)停止分泌的位置(據(jù)Fan et al., 2011)arrows show the locations of growth layers formed during low temperature in winter(after Fan et al., 2011)
ST5牡蠣殼體記錄的2002—2004年3年的冬季低溫季節(jié)平均最重δ18OV-PDB為 1.63‰, 夏季高溫季節(jié)平均最輕δ18OV-PDB為–3.41‰, 從冬季低溫季節(jié)到夏季高溫季節(jié)的平均變化幅度為5.04‰(表3)。
3 討論
Kirby等人(1998)對(duì)河口環(huán)境下自然生長(zhǎng)的現(xiàn)生活牡蠣(C. virginica)殼體與生長(zhǎng)環(huán)境之間的同位素平衡狀況進(jìn)行了分析研究。通過(guò)對(duì)活牡蠣生長(zhǎng)的水體溫度與鹽度的長(zhǎng)期監(jiān)測(cè)和一個(gè)航次內(nèi)采集的 8個(gè)水樣的氧同位素比值測(cè)定, 根據(jù)實(shí)測(cè)δ18O值與對(duì)應(yīng)的水體鹽度值, 擬合獲得了水體δ18O-鹽度線性關(guān)系式, 然后根據(jù)殼體的氧同位素分餾方程(Epstein et al., 1953)和實(shí)測(cè)水溫、水體氧同位素組成(由水體鹽度計(jì)算而來(lái)), 計(jì)算出在該水體中生長(zhǎng)的牡蠣分泌的殼體δ18O理論值, 對(duì)比分析結(jié)果顯示活牡蠣殼體與生長(zhǎng)水體之間的氧同位素完全達(dá)到平衡, 殼體在其生長(zhǎng)時(shí)刻真實(shí)地記錄了周圍水體的環(huán)境信息(Kirby et al., 1998)。
Epstein等人通過(guò)實(shí)驗(yàn)室可控溫度、鹽度水箱培養(yǎng)的軟體生物殼體的穩(wěn)定同位素比值分析, 發(fā)現(xiàn)殼體與存活水體之間的同位素達(dá)到平衡, 并且獲得了殼體氧同位素組成與水體氧同位素組成及水溫之間的定量關(guān)系式(Epstein et al., 1953)。
在殼體與生長(zhǎng)水體的氧同位素達(dá)到平衡的條件下, 若不考慮殼體生長(zhǎng)區(qū)水體鹽度變化對(duì)水體δ18O的貢獻(xiàn), 則殼體記錄的季節(jié)性δ18O變化則完全由水溫變化所致(程紅光等, 2009)。當(dāng)水溫升高 1°C, 貝殼的δ18OV-PDB則減小 0.24‰~0.25‰(Epstein et al.,1953), 因此, ST5記錄的 5.04‰季節(jié)性δ18OV-PDB變化幅度對(duì)應(yīng)的水溫變化幅度為 21~20.2℃。而 ST5殼體生長(zhǎng)區(qū)的實(shí)測(cè)水溫變化范圍為–0.7~27.1(℃表1), 變化幅度達(dá)27.8℃。研究區(qū)的夏季最高水溫低于Crassostrea屬牡蠣的熱休克溫度(28; Ke℃nnedy et al., 1996; Surge et al., 2001)。并且研究區(qū)的牡蠣殼體貝輪中也未見(jiàn)任何夏季高溫?zé)嵝菘擞涗?范昌福等,2010a; Fan et al., 2011)。因而, 殼體并未記錄水溫低于 6.5℃(6.1~6.9℃)的生長(zhǎng)環(huán)境信息, 即當(dāng)水溫低于6.5℃時(shí), 牡蠣停止分泌殼體。然而, 研究區(qū)的水體鹽度是變化的。當(dāng)考慮水體鹽度變化對(duì)水體δ18O的貢獻(xiàn)時(shí), 牡蠣停止分泌殼體的水溫將更低。
從冬季到夏季, 殼體生長(zhǎng)區(qū)的水體鹽度變化范圍為31.6×10-12~29.5×10-12(表1), 變化幅度為2.1×10-12。根據(jù)研究區(qū)水體鹽度與水體δ18O的變化關(guān)系δ18Owater,V-PDB/‰=0.12×S–26.7, 從冬季到夏季,殼體生長(zhǎng)區(qū)水體鹽度 2.1×10-12的變化對(duì)水體δ18OV-PDB變化的貢獻(xiàn)為 0.25‰。在考慮水體鹽度變化對(duì)水體δ18O的貢獻(xiàn)的情況下, ST5殼體記錄的水體溫度變化幅度為 22~21.2℃, 則牡蠣冬季停止分泌殼體的水溫約為5.5℃(5.1~5.9℃)。
根據(jù)渤海灣地區(qū)現(xiàn)生活牡蠣殼體的δ18O記錄和活牡蠣生長(zhǎng)水體的水溫與鹽度實(shí)測(cè)結(jié)果計(jì)算的牡蠣停止分泌殼體的最低平均水溫為 5.5℃, 這一水溫低于此前對(duì)同一地區(qū)的埋藏牡蠣殼體記錄的δ18O和文獻(xiàn)資料獲得的水文數(shù)據(jù)而計(jì)算出的 11.5℃的殼體停止分泌的最低水溫(Wang et al., 1995)。Ullmann等人(2010)對(duì)北海地區(qū)生長(zhǎng)的一個(gè)活牡蠣(C. gigas)殼體記錄的月分辨率δ18O和活牡蠣生長(zhǎng)水體的水溫與鹽度的監(jiān)測(cè)結(jié)果計(jì)算表明,C. gigas殼體停止分泌的水溫為 6℃。另有文獻(xiàn)報(bào)道,C. gigas可在水溫4~35℃的范圍內(nèi)生長(zhǎng), 它們甚至可以在–5℃的水溫環(huán)境下存活下來(lái)(Nehring, 2006)。
本文中的活牡蠣個(gè)體于2005年5月26日被采集時(shí), 已完成產(chǎn)卵并恢復(fù)快速生長(zhǎng)(圖4; Fan et al.,2011)。若C. gigas冬季停止分泌(春季恢復(fù)生長(zhǎng))的最低水溫為 11.5℃, 根據(jù)渤海灣地區(qū)的月均水溫資料, 牡蠣?wèi)?yīng)在4月份恢復(fù)生長(zhǎng)(表1), 至該個(gè)體被采集時(shí), 相隔僅1到2個(gè)月的時(shí)間, 而此時(shí)該個(gè)體不僅恢復(fù)了快速生長(zhǎng), 并且經(jīng)歷了春季產(chǎn)卵的緩慢生長(zhǎng)(范昌福等, 2010a; Fan et al., 2011)。據(jù)此實(shí)際情況,C. gigas恢復(fù)生長(zhǎng)的水溫應(yīng)更低, 渤海灣地區(qū)的現(xiàn)生ST5活牡蠣在平均水溫達(dá)5°C左右的3月份便恢復(fù)生長(zhǎng)。因此, 在進(jìn)一步獲得更準(zhǔn)確的數(shù)據(jù)結(jié)果之前, 采用5.5℃作為渤海灣地區(qū)C. gigas冬季停止分泌的最低水溫。
研究區(qū)的冬季最低水溫已低于 0℃, 遠(yuǎn)低于牡蠣分泌殼體的下限水溫, 因此, 一年之中低于下限水溫的季節(jié)(的信息)未被牡蠣殼體記錄。渤海灣地區(qū)牡蠣殼體的一個(gè)年生長(zhǎng)周期中最重的δ18O值分別紀(jì)錄了12月殼體停止分泌時(shí)刻和來(lái)年3月殼體恢復(fù)生長(zhǎng)時(shí)刻的生長(zhǎng)環(huán)境信息, 二者之間存在一個(gè)水溫低于5.5°C時(shí)出現(xiàn)的殼體分泌間斷期。
4 結(jié)論
渤海灣地區(qū)現(xiàn)生活牡蠣(C. gigas)冬季停止分泌/春季恢復(fù)生長(zhǎng)的最低水溫為 5.5℃, 該地區(qū)牡蠣生長(zhǎng)的季節(jié)為3月至12月。牡蠣殼體的一個(gè)年生長(zhǎng)周期中最重的δ18O值分別紀(jì)錄了 12月殼體停止分泌時(shí)刻和次年 3月殼體恢復(fù)生長(zhǎng)時(shí)刻的生長(zhǎng)環(huán)境信息。當(dāng)水溫低于5.5℃時(shí)(12月至次年3月之間), 牡蠣停止分泌殼體, 其生長(zhǎng)環(huán)境信息未能被記錄。
程紅光, 李心清, 袁洪林, 王克卓. 2009. 泥盆紀(jì)海水的碳、氧同位素變化——來(lái)自腕足化石的同位素記錄[J]. 地球?qū)W報(bào),30(1): 79-87.
范昌福, 高抒, 王宏. 2006. 渤海灣西北岸全新世埋藏牡蠣礁建造記錄中的間斷及其解釋[J]. 海洋地質(zhì)與第四紀(jì)地質(zhì),26(6): 27-35.
范昌福, 李建芬, 王宏, 閻玉忠, 王福, 裴艷東, 商志文. 2005a.渤海灣西北岸大吳莊牡蠣礁測(cè)年與古環(huán)境變化[J]. 地質(zhì)調(diào)查與研究, 28(2): 124-129.
范昌福, 王宏, 李建芬, 裴艷東, 王福, 商志文. 2005b. 渤海灣西北岸牡蠣礁體對(duì)區(qū)域性構(gòu)造活動(dòng)與水動(dòng)型海面變化的響應(yīng)[J]. 第四紀(jì)研究, 25(2): 235-244.
范昌福, 裴艷東, 王宏, 劉志廣, 王福, 商志文, 車?yán)^英, 田立柱.2007. 渤海灣西北岸埋藏牡蠣礁體中的殼體形態(tài)與沉積環(huán)境[J]. 第四紀(jì)研究, 27(5): 806-813.
范昌福, 王宏, 裴艷東, K?NIGER P, 李延河. 2010a. 牡蠣殼體的同位素貝殼年輪研究[J]. 地球科學(xué)進(jìn)展, 25(2): 163-173.
范昌福, 王宏, 裴艷東, 李延河. 2010b. 渤海灣西岸牡蠣殼體形態(tài)、生長(zhǎng)速率與生長(zhǎng)環(huán)境的關(guān)系[J]. 海洋通報(bào), 29(5):526-533.
范昌福, 王宏, 裴艷東, 李延河. 2010c. 穩(wěn)定同位素貝殼年輪指示的牡蠣礁體水平夾層成因[J]. 第四紀(jì)研究, 30(5):994-1002.
范昌福, 裴艷東, 田立柱, 王福, 劉志廣, 王宏. 2010d. 渤海灣西部淺海區(qū)活牡蠣礁調(diào)查結(jié)果及資源保護(hù)建議[J]. 地質(zhì)通報(bào), 29(5): 660-667.
范昌福, 王宏, 裴艷東, 劉志廣, 王福, 田立柱, 商志文. 2008.渤海灣西北岸濱海湖埋藏牡蠣礁古生態(tài)環(huán)境[J]. 海洋地質(zhì)與第四紀(jì)地質(zhì), 28(1): 33-41.
方晶, 范昌福, 王海峰, 白鐵磊, 商志文. 2011. 粘土混濁水電導(dǎo)率對(duì)牡蠣礁水平夾層成因的解釋——以渤海灣西北岸大吳莊牡蠣礁為例[J]. 第四紀(jì)研究, 31(4): 746-748.
耿秀山, 傅命左, 徐孝詩(shī), 李培英. 1991. 現(xiàn)代牡蠣礁發(fā)育與生態(tài)特征及古環(huán)境意義[J]. 中國(guó)科學(xué)(B輯), (8): 867-875.
李冬玲, 范昌福, 黃 玥 , 蔣輝, 王宏, 商志文. 2009. 渤海灣西北岸中全新世埋藏牡蠣礁的硅藻記錄及古環(huán)境意義[J]. 海洋通報(bào), 28(3): 22-28.
林防, 李鳳林, 李建芬, 張玉發(fā). 2004. 渤海灣西北岸全新世介形類組合特征及海進(jìn)海退旋回[J]. 地球?qū)W報(bào), 25(1): 53-58.
商志文, 范昌福, 李冬玲, 田立柱, 裴艷東, 王福, 王宏. 2010.硅藻組合指示的兩個(gè)牡蠣礁體生長(zhǎng)環(huán)境的差異[J]. 海洋地質(zhì)與第四紀(jì)地質(zhì), 30(5): 33-39.
天津市海岸帶和海涂資源綜合調(diào)查領(lǐng)導(dǎo)小組辦公室. 1987. 天津市海岸帶和海涂資源綜合調(diào)查報(bào)告[M]. 北京: 海洋出版社.
王海峰, 裴艷東, 劉會(huì)敏, 范昌福, 王宏. 2011. 渤海灣全新世牡蠣礁: 時(shí)空分布和海面變化標(biāo)志點(diǎn)[J]. 地質(zhì)通報(bào), 30(9):1396-1404.
王宏, 范昌福, 李建芬, 李鳳林, 閻玉忠, 王云生, 張金起, 張玉發(fā).2006. 渤海灣西北岸全新世牡蠣礁研究概述[J]. 地質(zhì)通報(bào),25(3): 1-17.
王宏, 張玉發(fā), 張金起, 李建芬, 李鳳林, VAN STRYDONCK M,HENDRIX V. 2000. 渤海灣西岸第二道貝殼堤的細(xì)分及其年齡序列[J]. 地球?qū)W報(bào), 21(3): 320-327.
王宏. 1996. 渤海灣全新世貝殼堤和牡蠣礁的古環(huán)境[J]. 第四紀(jì)研究, (1): 71-79.
王強(qiáng), 李秀文, 張志良, 李鳳林, 邵軍, 白德中, 尚學(xué)熙, 唐若魯.1991. 天津地區(qū)全新世牡蠣灘的古海洋學(xué)意義[J]. 海洋學(xué)報(bào), 13(3): 371-380.
薛春汀. 2003. 天 津?qū)幒涌h 俵 口牡蠣礁剖面與海面變化關(guān)系的討論[J]. 地理科學(xué), 23(1): 49-51.
張忍順. 2004. 江蘇小廟洪牡蠣礁的地貌-沉積特征. 海洋與湖沼, 35(1): 1-7.
張璽, 樓子庚. 1956. 中國(guó)牡蠣的研究[J]. 動(dòng)物學(xué)報(bào), 8(1): 65-93.
BRIGAUD B, PUCéAT E, PELLENARD P, VINCENT B,JOACHIMSKI M M. 2008. Climatic fluctuations and seasonality during the Late Jurassic (Oxfordian-Early Kimmeridgian)inferred fromδ18O of Paris Basin oyster shells[J]. Earth and Planetary Science Letters, 273(1-2): 58-67.
CARBOTTE S M, BELL R E, RYAN W B F, MCHUGH C,SLAGLE A, NITSCHE F, RUBENSTONE J. 2004. Environmental change and oyster colonization within the Hudson River estuary linked to Holocene climate[J]. Geo-marine Letters, 24(4): 212-224.
CHENG Hong-guang, LI Xin-qing, YUAN Hong-lin, WANG Ke-zhuo. 2009. Carbon and Oxygen Isotope Variations of Devonian Seawater: Isotopic Records from Brachiopod Shell[J]. Acta Geoscientica Sinica, 30(1): 79-87(in Chinese with English abstract).
COGNIE B, HAURE J, BARLLé L. 2006. Spatial distribution in a temperate coastal ecosystem of the wild stock of the farmed oysterCrassostrea gigas(Thunberg)[J]. Aquaculture, 259(1-4):249-259.
DIEDERICH S. 2005. Differential recruitment of introduced Pacific oysters and native mussels at the North Sea coast: Coexistence possible?[J]. Journal of Sea Research, 53(4): 269-281.EPSTEIN S, BUCHSBAUM R, LOWENSTAM H A, CUREY H C.1953. Revised carbonate-water isotopic temperature scale[J].Geological Society of America Bulletin, 64(11): 1315-1326.
FAN C, KOENIGER P, WANG H, FRECHEN M. 2011. Ligamental increments of the mid-Holocene Pacific oysterCrassostrea gigasare reliable independent proxies for seasonality in the western Bohai Sea, China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 299(3-4): 437-448.
FAN Chang-fu, GAO Shu, WANG Hong. 2006. Interpreting the hiatus in Holocene oyster reefs on the Northwest Coast of Bohai Bay[J]. Marine Geology & Quaternary Geology, 26(6):27-35(in Chinese with English abstract).
FAN Chang-fu, LI Jian-fen, WANG Hong, YAN Yu-zhong, WANG Fu, PEI Yan-dong, SHANG Zhi-wen. 2005a. Age and Paleoenvironmental Change of Dawuzhuang Oyster Reef on the Northwest Coast of Bohai Bay[J]. Geological Survey and Research, 28(2): 124-129 (in Chinese with English abstract).
FAN Chang-fu, WANG Hong, LI Jian-fen, PEI Yan-dong, WANG Fu, SHANG Zhi-wen. 2005b. Oyster reefs response to the regional neotectonics and eustatic sea-level, Northwest Coast of Bohai Bay[J]. Quaternary Sciences, 25(2): 235-244 (in Chinese with English abstract).
FAN Chang-fu, PEI Yan-dong, WANG Hong, LIU Zhi-guang,WANG Fu, SHANG Zhi-wen, CHE Ji-ying, TIAN Li-zhu.2007. Correlation of oyster shell form and sediment environment from two buried oyster reefs on the Northwest Coast of Bohai Bay[J]. Quaternary Sciences, 27(5): 806-813 (in Chinese with English abstract).
FAN Chang-fu, WANG Hong, PEI Yan-dong, K?NIGER P, LI Yan-he. 2010a. Stable isotope sclerochronology study of oyster shells[J]. Advances in Earth Science, 25(2): 163-173 (in Chinese with English abstract).
FAN Chang-fu, WANG Hong, PEI Yan-dong, LI Yan-he. 2010b.Relationship among the form, growth rate and living environment of oyster shells in west coast of Bohai Bay[J]. Marine Science Bulletin, 29(5): 526-533(in Chinese with English abstract).
FAN Chang-fu, WANG Hong, PEI Yan-dong, LI Yan-he. 2010c.Origins of horizontal oyster interlayers in buried reefs revealed by stable isotope sclerochronology[J]. Quaternary Sciences, 30(5): 994-1002(in Chinese with English abstract).
FAN Chang-fu, PEI Yan-dong, TIAN Li-zhu, WANG Fu, LIU Zhi-guang, WANG Hong. 2010d. Living oyster reefs and its resource conservation in western Bohai Bay, China[J]. Geological Bulletin of China, 29(5): 660-667(in Chinese with English abstract).
FAN Chang-fu, WANG Hong, PEI Yan-dong, LIU Zhi-guang,WANG Fu, TIAN Li-zhu, SHANG Zhi-wen. 2008. Palaeoecological environment revealed by the buried binhaihu oyster reef on the northwest coast of Bohai Bay[J]. Marine Geology & Quaternary Geology, 28(1): 33-41(in Chinese with English abstract).
FANG Jing, FANG Chang-fu, WANG Hai-feng, BAI Tie-lei,SHANG Zhi-wen. 2011. A causal explanation for the intercalated horizontal layers of Dawuzhuang oyster reef by the clay-water electric conductivity[J]. Quaternary Sciences, 31(4):746-748(in Chinese with English abstract).
GENG Xiu-shan, FU Ming-zuo, XU Xiao-shi, LI Pei-ying. 1991.Living oyster reef development and ecological characteristics,and its palaeoenvironmental significance[J]. Science in China(Series B), (8): 867-875(in Chinese).
GONFIANTINI R, STICHLER W, ROZANSKI K. 1995. Standards and Intercomparison Materials Distributed by the International Atomic Energy Agency for Stable Isotope Measurements[C]//Reference and Intercomparison Materials for Stable Isotopes of Light elements, edited by the Isotope Hydrology Section of the International Atomic Energy Agency. IAEA,Vienna, Austria, 13-29.
KENNEDY V S, NEWELL R I E, EBLE A F. 1996. The Eastern oyster:Crassostrea virginica[M]. College Park, Maryland:Maryland Sea Grant Press.
KIRBY M X, SONIAT T M, SPERO H J. 1998. Stable isotope sclerochronology of Pleistocene and recent oyster shells(Crassostrea virginica)[J]. PALAIOS, 13(6): 560-569.
Leading Group Office of the Tianjin Comprehensive Survey of Coastal Zones and Shoals Resources. 1987. Report of Tianjin coastal zones and shoals resources survey[M]. Beijing: Ocean Press(in Chinese).
LI Dong-ling, FAN Chang-fu, HUANG Yue, JIANG Hui, WANG Hong, SHANG Zhi-wen. 2009. Paleo-environmental Reconstruction with Diatom Assemblages in the Buried Oyster Reef on the Northwest Coast of the Bohai Bay During the Middle Holocene[J]. Marine Science Bulletin, 28(3): 22-28(in Chinese with English abstract).
LIN Fang, LI Feng-lin, LI Jian-fen, ZHANG Yu-fa. 2004. Holocene Ostracoda Assemblages and Marine Transgression-Regression Cycle in the Northwestern Coastal Area of the Bohai Gulf[J].Acta Geoscientica Sinica, 25(1): 53-58(in Chinese with English abstract).
NEHRING S. 2006. NOBANIS-Invasive Alien Species Fact Sheet-Crassostrea gigas. From: Online database of the North European and Baltic Network on Invasive Alien Species-NOBANIS[DB/OL]. [2011-11-14]. www.nobanis.org/files/factsheets/Crassostrea_gigas.pdf.
RYE D M, SOMMER M A. 1980. Reconstructing paleotemperature and paleosalinity regimes with oxygen isotope[M]//RHODES D C, LUTZ R A. Skeletal Growth of Aquatic Organisms. New York: Plenum Press: 169-201.
SCH?NE B R, FIEBIG J, PFEIFFER M, GLEΒ R, HICKSON J,JOHNSON L A A, DREYER W, OSCHMANN W. 2005.Climate records from a bivalved Methuselah (Arctica islandica, Mollusca; Iceland)[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 228(1-2): 130-148.
SCH?NE B R, FREYRE CASTRO A D, FIEBIG J, HOUK S D,OSCHMANN W, KR?NCKE I. 2004. Sea surface water temperatures over the period 1884-1983 reconstructed from oxygen isotope ratios of a bivalve mollusk shell (Arctica islandica,southern North Sea)[J]. Palaeogeography, Palaeoclimatology,Palaeoecology, 212: 215-232.
SHANG Zhi-wen, FAN Chang-fu, LI Dong-lin, TIAN Li-zhu, PEI Yan-dong, WANG Fu, WANG Hong. 2010. Revealing paleo-environment difference of two oyster reefs by diatom assemblages in the northwest coast of Bohai Bay[J]. Marine Geology & Quaternary Geology, 30(5): 33-39(in Chinese with English abstract).
SIMSTICH J, HARMS I, KARCHER M J, ERLENKEUSER H,STANOVOY V, KODINA L, BAUCH D, SPIELHAGEN R F.2005. Recent freshening in the Kara Sea (Siberia) recorded by stable isotopes in Arctic bivalve shells[J]. Journal of Geophysical Research, 110: C08006.
SURGE D, LOHMANN K C, DETTMAN D L. 2001. Controls on isotopic chemistry of the American oyster,Crassostrea virginica: implications for growth patterns[J]. Palaeogeography,Palaeoclimatology, Palaeoecology, 172(3-4): 283-296.
TCHANG Si, LOU Tze-kong. 1956. A study on Chinese oysters[J].Acta Zoologica Sinica, 8(1): 65-93(in Chinese with English abstract).
THUNBERG C P. 1793. Tekning och Beskrifning pa en stor Ostronsort ifran Japan[J]. Kongliga Vetenskaps Academiens Nya Handlingar, 14(4-6): 140-142
ULLMANN C V, WIECHERT U, KORTE C. 2010. Oxygen isotope fluctuations in a modern North Sea oyster (Crassostrea gigas)compared with annual variations in seawater temperature: Implications for palaeoclimate studies[J]. Chemical Geology,277(1): 160-166.
WANG H, KEPPENS E, NIELSEN P, VAN RIET A. 1995. Oxygen and carbon isotope study of the Holocene oyster reefs and paleoenvironmental reconstruction on the Northwest Coast of Bohai Bay, China[J]. Marine Geology, 124(1-4): 289-302.
WANG Hai-feng, PEI Yan-dong, LIU Hui-min, FAN Chang-fu,WANG Hong. 2011. Holocene oyster reefs: spatial and temporal distribution and sea level in dicators in Bohai Bay[J].Geological Bulletin of China, 30(9): 1396-1404(in Chinese with English abstract).
WANG Hong, FAN Chang-fu, LI Jian-fen, LI Feng-lin, YAN Yu-zhong, WANG Yun-sheng, ZHANG Jin-qi, ZHANG Yu-fa.2006. Holocene oyster reefs on the northwest coast of the Bohai Bay, China[J]. Geological Bulletin of China, 25(3):1-17(in Chinese with English abstract).
WANG Hong, ZHANG Yu-fa, ZHANG Jin-qi, LI Jian-fen, LI Feng-lin, VAN STRYDONCK M, HENDRIX V. 2000. The Chenier II on the Western Coast of Bohai Bay: Its Subdivision and Amended Time-framework[J]. Acta Geoscientica Sinica,21(3): 320-327(in Chinese with English abstract).
WANG Hong. 1996. Palaeoenvironment of the Holocene Cheniers and oyster reefs in the Bohai bay, China[J]. Quaternary Sciences, (1): 71-79(in Chinese with English abstract).
WANG Qiang, LI Xiu-wen, ZHANG Zhi-liang, LI Feng-lin, SHAO Jun, BAI De-zhong, SHANG Xue-xi, TANG Ruo-lu. 1991.Palaeo-oceanological significance in Holocene ostracean banks in Tianjin area[J]. Acta Oceanologica Sinica, 13(3):371-380(in Chinese with English abstract).
WISSHAK M, CORREA M L, GOFAS S, SALAS C, TAVIANI M,JAKOBSEN J, FREIWALD A. 2009. Shell architecture, element composition, and stable isotope signature of the giant deep-sea oysterNeopycnodonte zibrowiisp. n. from the NE Atlantic[J]. Deep Sea Research Part I: Oceanographic Research Papers, 56(3): 374-407.
XUE Chun-ting. 2003. Discussion on relationship between Holocene sea level fluctuation and Biaokou Section, Ninghe, Tianjin, China[J]. Scientia Geographica Sinica, 23(1): 49-51(in Chinese with English abstract).
ZHANG Jia-fu, FAN Chang-fu, WANG Hong, ZHOU Li-ping.2007. Chronology of an oyster reef on the coast of Bohai Bay,China: Constraints from optical dating using different luminescence signals from fine quartz and polymineral fine grains of coastal sediments[J]. Quaternary Geochronology,2(1-4): 71-76.
ZHANG Ren-shun. 2004. The geomorphology-sedimentology character of oyster reef in Xiaomiaohong tidal channel, Jiangsu Province[J]. Oceanologia et Limnologia Sinica, 35(1):1-7(in Chinese with English abstract).
致謝: 中國(guó)地質(zhì)科學(xué)院礦產(chǎn)資源研究所李延河研究員給予了指導(dǎo); 德國(guó)漢諾威萊布尼茲應(yīng)用地質(zhì)科學(xué)研究所Manfred Frechen教授、Paul K?niger博士和Astrid Jaeckel女士幫助測(cè)定了本文中的殼體同位素樣品; 南京大學(xué)內(nèi)生金屬礦床成礦機(jī)制研究國(guó)家重點(diǎn)實(shí)驗(yàn)室楊濤博士幫助測(cè)定了水樣氧、碳同位素;中國(guó)地質(zhì)調(diào)查局天津地質(zhì)調(diào)查中心李建芬博士、王福博士、田立柱博士、商志文博士和耿巖碩士參加了野外樣品的采集和貝殼年輪研究樣品的前處理工作, 在此一并致謝!
The Lowest Sea Surface Temperatures for Stopping Secretion in Winter and Resuming Growth in Spring Recorded by the Living Pacific Oyster (Crassostrea gigas) Shell in the Bohai Bay
FAN Chang-fu1,2), WANG Hong3), PEI Yan-dong3), WANG Hai-feng3)1)Institute of Mineral Resources, Chinese Academy of Geological Sciences, Beijing100037;2)MRL Key Laboratory of Metallogeny and Mineral Assessment, Institute of Mineral Resources,Chinese Academy of Geological Sciences, Beijing100037;3)Tianjin Center, China Geological Survey, Tianjin 300170
Oyster reefs are widely distributed along coastal and estuary areas. The in situ living oyster shell is a good object for studying and reconstructing the climate and environment change. With a living Pacific oyster(Crassostrea gigas) shell collected from the northwest coast of Bohai Bay as the study object and according to the analytical results of stable isotope of O in the continuous growth micro-layers of the shell as well as monthly average sea surface salinity (SSS) and sea surface temperature (SST), the authors obtained the lowest sea surface temperature recorded by theC. gigasshell in winter. In Bohai Bay area, the cease of shell growth ofC.gigaslasts for up to 3 months per year from December to March, when the water temperature decreases to 5.5.℃
Mollusca shell; stopping secretion; oxygen isotope; SST; Bohai Bay
Q915.817; P597.2
A
10.3975/cagsb.2012.06.15
本文由國(guó)家自然科學(xué)基金項(xiàng)目(編號(hào): 40872106)資助。
2012-08-29; 改回日期: 2012-10-21。責(zé)任編輯: 張改俠。
范昌福, 男, 1979年生。副研究員。主要從事同位素地球化學(xué)研究。E-mail: tjfchangfu@163.com。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
