天山凍土中產(chǎn)低溫蛋白酶菌株的篩選、鑒定及酶學(xué)性質(zhì)*
2012-09-12 13:21:58祁丹丹楊瑞金華霄沈蓮蓮張文斌趙偉
食品與發(fā)酵工業(yè) 2012年7期
關(guān)鍵詞:生長
祁丹丹,楊瑞金,華霄,沈蓮蓮,張文斌,趙偉
1(江南大學(xué)食品科學(xué)與技術(shù)國家重點(diǎn)實(shí)驗(yàn)室,江蘇無錫,214122)
2(江南大學(xué)食品學(xué)院,江蘇 無 錫,214122)
天山凍土中產(chǎn)低溫蛋白酶菌株的篩選、鑒定及酶學(xué)性質(zhì)*
祁丹丹1,楊瑞金2,華霄2,沈蓮蓮1,張文斌2,趙偉2
1(江南大學(xué)食品科學(xué)與技術(shù)國家重點(diǎn)實(shí)驗(yàn)室,江蘇無錫,214122)
2(江南大學(xué)食品學(xué)院,江蘇 無 錫,214122)
借助形態(tài)觀察、生理生化指標(biāo)測(cè)定及16S rDNA序列分析,對(duì)從新疆天山凍土中篩選得到的產(chǎn)低溫蛋白酶菌株進(jìn)行鑒定,確定其為Pseudomonas sp.(假單胞菌屬)N-8。通過單因素試驗(yàn)和正交實(shí)驗(yàn)對(duì)其發(fā)酵條件進(jìn)行優(yōu)化,確定最佳發(fā)酵培養(yǎng)基配方為(g/L):葡萄糖10,酵母粉8,酪蛋白20,K+0.4,pH 7.0。酶學(xué)性質(zhì)研究表明,該菌產(chǎn)生的低溫蛋白酶最適作用pH為7.0,在pH 7.0~8.0呈現(xiàn)良好的穩(wěn)定性;最適作用溫度為25℃,在25、35、45℃下均呈現(xiàn)良好的穩(wěn)定性;金屬離子Cu2+、K+、Ca2+、Mn2+對(duì)酶活有一定的抑制作用,其中Cu2+抑制作用明顯,F(xiàn)e2+、Mg2+、Na+、Zn2+對(duì)酶活力有不同程度的激活作用。
耐冷菌,低溫蛋白酶,酶學(xué)性質(zhì),假單胞菌屬
天山凍土屬于我國高海拔高山常年凍土,凍土環(huán)境中微生物會(huì)產(chǎn)生一系列適應(yīng)低溫、寡營養(yǎng)、強(qiáng)輻射、凍融等極端因子的分子機(jī)制,為低溫微生物生理多樣性提供了可能[1]。低溫酶由于在低溫環(huán)境下具有較高的酶活性,有助于在保證催化效率的前提下降低反應(yīng)溫度和縮短反應(yīng)時(shí)間,從而大幅度節(jié)約能源[2],因而在食品加工、日化產(chǎn)品、皮革加工等需要低溫催化的行業(yè)中具有較高的開發(fā)利用價(jià)值[3]。目前發(fā)現(xiàn),產(chǎn)低溫蛋白酶的菌有假單胞菌屬(Psedomonas)[4]、黃桿菌屬(Flavobacterium)[5]、氣單孢菌屬(Aeromonas)[6]、希瓦氏菌屬(Shewanella)[7]、弧菌屬(Vibrio)[8]。這些低溫菌在永久性低溫環(huán)境下經(jīng)過長期的進(jìn)化適應(yīng),已有著適應(yīng)低溫環(huán)境的特殊結(jié)構(gòu)與生理生化機(jī)制。它們所產(chǎn)生的低溫蛋白酶也有著獨(dú)特的生理生化特征與結(jié)構(gòu)特征。有關(guān)低溫微生物的資源勘探與代謝活性產(chǎn)物研究,已成為國際微生物學(xué)研究的熱點(diǎn)之一[9],我國的一些研究機(jī)構(gòu)也積極開展低溫蛋白酶生產(chǎn)菌株的篩選工作[10-11]。
本課題組從新疆凍土中篩選出1株產(chǎn)低溫蛋白酶的耐冷菌株,經(jīng)鑒定為假單胞菌屬,對(duì)其發(fā)酵條件進(jìn)行優(yōu)化以提高其產(chǎn)酶能力,并對(duì)其所產(chǎn)低溫蛋白酶的粗酶酶學(xué)性質(zhì)進(jìn)行了初步研究,為極地微生物產(chǎn)低溫蛋白酶的研究打下一定的基礎(chǔ)。
1 材料和方法
1.1 實(shí)驗(yàn)材料
樣品采自新疆天山冰川海拔3845 m處。沿2 m深的凍土剖面每隔20 cm取樣,迅速將其裝入已滅菌的保鮮盒內(nèi),置于車載冰箱保存,運(yùn)回實(shí)驗(yàn)室后,在超凈臺(tái)上削去表皮可能受到污染的樣品,于-20℃保存?zhèn)溆谩?/p>
1.2 培養(yǎng)基
PYG富集培養(yǎng)基(g/L):蛋白胨0.5,酵母粉1,葡萄糖1,CaCl20.02,K2HPO4·3H2O 0.1,MgSO4·7H2O 0.04,NaCl 0.5,pH 7.2;
初篩培養(yǎng)基(g/L):蛋白胨0.05,酵母膏0.03,酪蛋白1,瓊脂1.5,pH 7.0;
發(fā)酵培養(yǎng)基(g/L):葡萄糖1,酵母粉0.4,酪蛋白1,MgSO4·7H2O 0.02,K2HPO4·3H2O 0.1,pH 7.0。
1.3 產(chǎn)低溫蛋白酶菌種的篩選
初篩:吸取富集后的培養(yǎng)液用倍比稀釋法制不同的稀釋度移入初篩平板中,涂勻,15℃培養(yǎng)。待菌落長好后,挑選透明圈較大菌株,斜面保存。
溫度復(fù)篩:以40℃為上限生長溫度[12],即在該溫度不生長的菌株為耐冷菌。將初篩得到的產(chǎn)蛋白酶菌株在初篩平板上點(diǎn)樣,在40℃不生長的菌株,為試驗(yàn)菌株。
蛋白酶活力的測(cè)定:按照SB/T10317—1999規(guī)定的福林(Folin)試劑顯色法測(cè)定酶活,進(jìn)一步篩選出產(chǎn)酶能力強(qiáng)的菌株。底物2%的酪蛋白,pH 7.2磷酸鹽緩沖液,15℃反應(yīng)10 min,660 nm處測(cè)定OD值,計(jì)算酶活力。在上述條件下,1 mL酶液每分鐘反應(yīng)產(chǎn)生1 μg酪氨酸為1個(gè)酶活單位(U)。
1.4 菌種鑒定
1.4.1 菌落形態(tài)觀察、生理生化試驗(yàn)
將所得菌株進(jìn)行革蘭氏染色,在電鏡下觀察菌株形態(tài),參考《伯杰細(xì)菌鑒定手冊(cè)》(第八版)[13]及《常見細(xì)菌系統(tǒng)鑒定手冊(cè)》[14]進(jìn)行生理生化鑒定。
1.4.216S rDNA PCR擴(kuò)增和序列分析
擴(kuò)增引物為通用引物,
上游引物(F):5'-AGAGTTTGATCCTGGCTCAG-3',
下游引物(R):5'-TACGGCTACCTTGTTACGACTT-3'。
PCR反應(yīng)過程:94℃預(yù)變性4 min,94℃變性1 min,55℃退火1 min,72℃延伸2 min,35個(gè)循環(huán);72℃補(bǔ)齊10 min。用標(biāo)準(zhǔn)PCR反應(yīng)體系進(jìn)行PCR擴(kuò)增。PCR產(chǎn)物電泳(1.0%瓊脂糖凝膠),回收目的片段,純化產(chǎn)物連接pMD18-T載體,轉(zhuǎn)入感受態(tài)E.coli DH5α,LB/Amp/X-Gal/IPTG平板上挑選白斑,抽提質(zhì)粒,PCR鑒定陽性重組菌,送上海博尚生物技術(shù)有限公司測(cè)序。
1.4.3 系統(tǒng)發(fā)育樹分析
利用CLUSTAL X將所測(cè)序列與Genbank中核酸序列進(jìn)行比對(duì),選用MEGA 5.0軟件Kimura 2-parameter距離模型進(jìn)行neighbour-joining分析生成系統(tǒng)發(fā)育樹,并進(jìn)行分析。
1.5 發(fā)酵條件優(yōu)化
1.5.1 培養(yǎng)時(shí)間對(duì)菌體生長及產(chǎn)酶的影響
連續(xù)培養(yǎng)72 h,每隔6 h取1次樣,測(cè)菌體生長量(OD600nm),同時(shí)制備粗酶液,測(cè)定酶活。
1.5.2 發(fā)酵溫度對(duì)菌體生長及產(chǎn)酶的影響
分別在10、15、20、25、30、35℃下進(jìn)行搖瓶,考察不同發(fā)酵溫度對(duì)菌體生長和產(chǎn)酶的影響。
1.5.3 發(fā)酵培養(yǎng)基組成對(duì)產(chǎn)酶的影響
選取葡萄糖、酵母粉、酪蛋白、K+4個(gè)因子,采用4因素3水平正交表L9(34)設(shè)計(jì)正交實(shí)驗(yàn),考察培養(yǎng)基組成對(duì)產(chǎn)酶的影響。
1.5.4 發(fā)酵條件對(duì)產(chǎn)酶的影響
選取種齡、起始pH、接種量、裝液量4個(gè)因子,采用正交表L9(34)設(shè)計(jì)正交實(shí)驗(yàn),考察不同發(fā)酵條件對(duì)產(chǎn)酶的影響。
1.6 粗酶酶學(xué)性質(zhì)的研究
1.6.1 粗酶的最適溫度和熱穩(wěn)定性
最適溫度測(cè)定:將酶活力測(cè)定中的反應(yīng)溫度分別控制在10、15、20、25、30、35、40、45、50、55、65℃,其他條件不變,分別測(cè)定酶活。
熱穩(wěn)定性測(cè)定:將粗酶液分別置于25、35、45、55℃,保溫15、30、45、60 min,冷卻后測(cè)酶活。
1.6.2 粗酶的最適pH和pH穩(wěn)定性
最適作用pH測(cè)定:配置不同pH的緩沖液,并用相應(yīng)緩沖液配置2%酪蛋白溶液作底物,測(cè)定酶活。
pH穩(wěn)定性測(cè)定:將用不同pH緩沖液處理的酶液分別放置在25℃下保溫1 h后測(cè)酶活。其中pH 5.0~7.0用磷酸鹽緩沖液,pH 8.0~9.0用Tris-HCl緩沖液,pH 10.0~11.0用硼砂-氫氧化鈉緩沖液。
1.6.3 金屬離子對(duì)酶活的影響
不同金屬離子Fe2+、Mn2+、K+、Na+、Cu2+、Ca2+、Mg2+、Zn2+水溶液與等體積蛋白酶混合,使最終離子濃度為5 mmol/L,25℃反應(yīng)15 min,測(cè)酶活性。
2 結(jié)果與討論
2.1 產(chǎn)低溫蛋白酶菌株的篩選
從天山凍土中通過酪蛋白平板法篩選到66株具有蛋白酶活性菌株,經(jīng)溫度復(fù)篩及Folin法測(cè)定酶活,篩選得到5株產(chǎn)蛋白酶活力較高的耐冷菌株,結(jié)果如表1所示。由表1可以看出,菌株N-8所產(chǎn)蛋白酶酶活最高,選取該菌做進(jìn)一步研究。
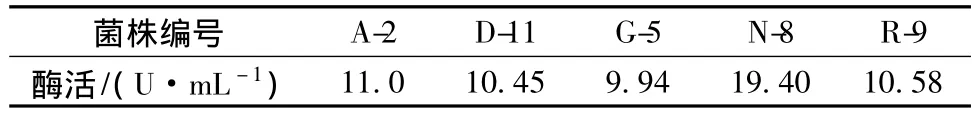
表1 產(chǎn)蛋白酶菌株篩選結(jié)果
2.2 菌種鑒定
2.2.1 菌落形態(tài)
該菌為革蘭氏陰性菌,菌體呈桿狀,細(xì)胞單個(gè),大小為(0.5~0.8)μm×(2.0~3.2)μm,有莢膜,有鞭毛(見圖1);在酪蛋白平板上,菌落直徑3~4 mm,呈圓形,乳白色,不透明,表面光滑,邊緣整齊,易挑取,產(chǎn)生明顯的透明圈。
2.2.2 生理生化特性
由表2生理生化特性可以發(fā)現(xiàn),該菌株接觸酶、精氨酸雙水解酶、硝酸鹽(還原)反應(yīng)為陽性,能水解明膠,硫化氫試驗(yàn)、伏-普二氏試驗(yàn)(VP)、甲基紅試驗(yàn)(MR)為陰性,不能發(fā)酵葡萄糖、乳糖、麥芽糖、蔗糖、纖維二糖、鼠李糖、棉子糖。
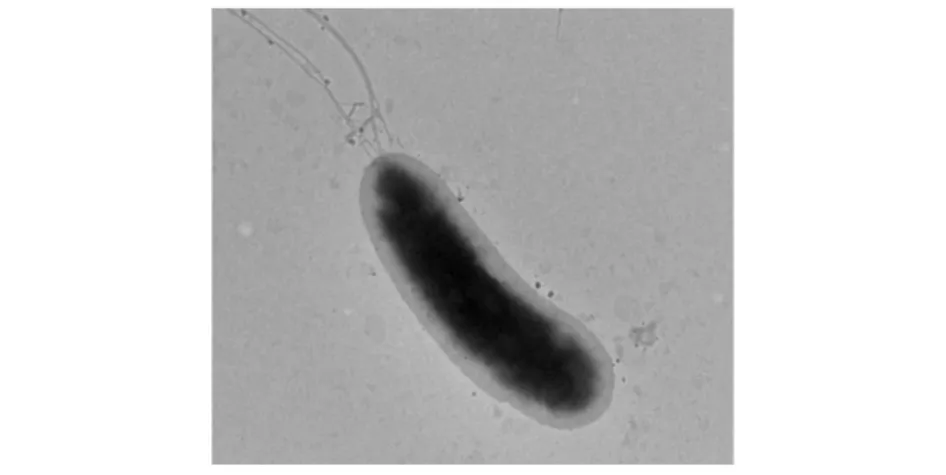
圖1 N-8菌株透射電鏡照片(5000×8)

表2 分離菌株的生理生化特性
2.2.3 系統(tǒng)發(fā)育樹分析
經(jīng)PCR擴(kuò)增后,該菌株16S rDNA片段長約1500 bp,將其16S rDNA序列同NCBI數(shù)據(jù)庫的相似序列進(jìn)行同源性比較,如圖2所示。Blast分析顯示該菌與Pseudomonas extremaustralis 14-3在同一分支,二者序列相似性為99%。綜合菌株的形態(tài)特征、生理生化特性、16S rDNA序列以及系統(tǒng)進(jìn)化樹的結(jié)果,可以將分離菌株歸為Pseudomonas屬,命名為Pseudomonas sp.N-8。
2.3 發(fā)酵條件優(yōu)化
2.3.1 培養(yǎng)時(shí)間對(duì)菌體生長及產(chǎn)酶的影響
圖3顯示了培養(yǎng)時(shí)間對(duì)菌體產(chǎn)酶的影響。在對(duì)數(shù)生長期,雖然菌體生長旺盛,但酶活很低。隨著穩(wěn)定期到來,酶活迅速升高。該菌在48 h酶活最高,隨著生長速度減慢,酶活也逐漸降低。從細(xì)菌生長曲線分析,酶的積累主要發(fā)生在對(duì)數(shù)生長后期,說明對(duì)數(shù)生長后期菌體不再大量繁殖,主要產(chǎn)生蛋白酶。因此酶的收獲應(yīng)控制在細(xì)菌生長穩(wěn)定期。
2.3.2 培養(yǎng)溫度對(duì)菌體生長及產(chǎn)酶的影響

圖2 菌株P(guān)seudomonas sp.N-8及相關(guān)細(xì)菌16S rDNA序列系統(tǒng)發(fā)育樹

圖3 培養(yǎng)時(shí)間對(duì)菌體產(chǎn)酶的影響
圖4 顯示培養(yǎng)溫度對(duì)菌體產(chǎn)酶的影響。菌株在20℃菌體生長及產(chǎn)酶情況較好,溫度高于或低于此溫度菌體生長和產(chǎn)酶能力有明顯下降的趨勢(shì)。

圖4 培養(yǎng)溫度對(duì)菌體產(chǎn)酶的影響
2.3.3 發(fā)酵培養(yǎng)基組成對(duì)產(chǎn)酶的影響
極差分析結(jié)果表明(表3),各因素對(duì)產(chǎn)酶的影響大小順序?yàn)?酪蛋白>酵母粉>葡萄糖>K+;最佳培養(yǎng)基配方為(g/L):葡萄糖10,酵母粉8,酪蛋白20,K+0.4;按最佳組合測(cè)定發(fā)酵液酶活為47.34 U/mL。
2.3.4 發(fā)酵條件對(duì)產(chǎn)酶的影響
極差分析結(jié)果表明(表4),各因素對(duì)產(chǎn)酶的影響大小順序?yàn)?種齡>起始pH>接種量>裝液量;最佳培養(yǎng)條件為:種齡24 h,起始pH 7,接種量4%,裝液量50 mL/250 mL。按最佳組合測(cè)定發(fā)酵液酶活為49.56 U/mL。

表3 培養(yǎng)基組成的正交試驗(yàn)設(shè)計(jì)與結(jié)果

表4 發(fā)酵條件的正交試驗(yàn)設(shè)計(jì)與結(jié)果
2.4 粗酶酶學(xué)性質(zhì)研究
2.4.1 粗酶的最適溫度
溫度對(duì)Pseudomonas sp.N-8產(chǎn)低溫蛋白酶活力的影響結(jié)果如圖5所示。該酶在10℃時(shí)可保持約60%的相對(duì)酶活,隨著溫度的升高,酶活逐漸提高,在25℃酶活最高,高于此溫度酶活開始下降。在溫度20~30℃范圍內(nèi),低溫蛋白酶都能保持較高的酶活,溫度超過40℃,酶活急劇下降,65℃時(shí)酶活不足最高酶活的10%。因此,N-8所產(chǎn)蛋白酶的最適溫度為25℃。
2.4.2 熱穩(wěn)定性

圖5 酶的最適溫度
菌株N-8所產(chǎn)低溫蛋白酶熱穩(wěn)定性如圖6所示。該酶在25℃呈現(xiàn)良好的穩(wěn)定性,保溫60 min仍可保持95%的酶活;在35、45℃保溫45 min酶活喪失了20%左右;在55℃下保溫30 min后酶活喪失了75%。這與報(bào)道的低溫蛋白酶熱穩(wěn)定性相符,即在較高溫度下(>50℃)酶可快速失活[2,3]。

圖6 酶的熱穩(wěn)定性
2.4.3 粗酶的最適pH
pH對(duì)Pseudomonas sp.N-8產(chǎn)低溫蛋白酶活性的影響結(jié)果如圖7所示。菌株N-8所產(chǎn)低溫蛋白酶最適pH為7.0,在pH 7.0~7.5范圍內(nèi)保持較高酶活。在酸性或堿性環(huán)境中,酶活大幅度下降。故菌株N-8所產(chǎn)低溫蛋白酶為中性蛋白酶。

圖7 酶的最適pH值
2.4.4 pH的穩(wěn)定性
菌株N-8所產(chǎn)低溫蛋白酶pH穩(wěn)定性如圖8所示。菌株N-8所產(chǎn)低溫蛋白酶pH在7.0~8.0范圍內(nèi)相對(duì)酶活達(dá)到90%以上,在pH 5.0時(shí)酶活下降約40%,在pH 10.0時(shí)酶活下降約25%。說明菌株N-8所產(chǎn)低溫蛋白酶在pH 7.0~8.0范圍內(nèi)呈現(xiàn)較好的穩(wěn)定性。

圖8 酶的pH穩(wěn)定性
2.4.5 金屬離子對(duì)酶活的影響
金屬離子對(duì)Pseudomonas sp.N-8產(chǎn)低溫蛋白酶活性的影響結(jié)果如圖9所示。K+、Cu2+、Ca2+、Mn2+對(duì)蛋白酶有不同程度的抑制作用,其中Cu2+的抑制作用明顯;Fe2+、Mg2+、Na+、Zn2+對(duì)蛋白酶有不同程度的激活作用。

圖9 金屬離子對(duì)酶活的影響
3 結(jié)語
低溫蛋白酶主要由生存在低溫冷凍環(huán)境中的微生物產(chǎn)生,目前大多數(shù)產(chǎn)酶菌株是從南、北極和常年恒定在低溫的極端環(huán)境中分離而得。本課題從新疆天山長年冷凍的土壤中分離獲得1株產(chǎn)低溫蛋白酶的菌株,經(jīng)鑒定為Psedomonas屬,并對(duì)其生長、產(chǎn)酶條件和粗酶性質(zhì)做了初步研究。報(bào)道低溫蛋白酶的最適酶活溫度大部分為15~40℃,如史勁松等報(bào)道耐冷菌株SYP-A2-3所產(chǎn)低溫蛋白酶的最適酶活溫度為15℃[15],徐國英等報(bào)道Pseudoalterom onas sp.QI-1(假交替單胞菌屬)所產(chǎn)蛋白酶的最適酶活溫度為40℃[16]。本研究分離菌株產(chǎn)低溫蛋白酶的最適溫度為20℃,酶活最適作用溫度為25℃,55℃保持30 min酶活喪失75%,65℃下作用酶活僅保留不到10%。本文篩選的菌株為今后低溫酶的嗜冷機(jī)制研究及其在食品、洗滌劑、化妝品、皮革等工業(yè)上的應(yīng)用研究奠定基礎(chǔ)。
[1]張明,顧燕玲,徐宇麗,等.天山1號(hào)冰川底部沉積層產(chǎn)-半乳糖苷酶低溫菌株的系統(tǒng)發(fā)育分析及生理多樣性[J].微生物學(xué)報(bào),2011,51(12):1605-1615.
[2]Margesin R,Dieplinger H,Hofmann J,et a1.A cold-active extracellular metalloprotease from Pedobacter cryoconitis:production and properties[J].Research in Microbiology,2005,156(4):499-505.
[3]Quanfu W,Yanhua H.Purification and properties of an extracellular cold-active protease from the psychrophilic bacterium Pseudoalteromonas sp.NJ276[J].Biochemical Engineering Journal,2008,38(3):362-368.
[4]Rahman R N,Geok L P,Basri M.An organic solvent-stable alkaline protease from Pseudomonas aeruginosa strain K:Enzyme purification and characterization[J].Enzyme and Microbial Technology,2006,39(7):1484-1491.
[5]王躍軍,孫謐,洪義國.黃海黃桿菌YS-9412-130低溫堿性蛋白酶的制備工藝研究[J].水產(chǎn)研究,2001,22(2):11-19.
[6]Nakasone N,Toma C,Song T,et al.Purification and characterization of a novel metalloprotease isolated from Aeromonas caviae[J].FEMS Microbiology Letters,2004,207(1):127-132.
[7]Kulakova L,Galkin A,Nakayama T,et al.Improvement of thermostability of cold-active serine alkaline protease from the psychrotrophic bacterium Shewanella sp.strain Ac10 by rational mutagenesis[J].Journal of Molecular Catalysis B:Enzymatic,2003,22(1/2):113-117.
[8]Venugopal M,Saramma A V.Characterization of alkaline protease from Vibrio fluvialis strain VM10 isolated from a mangrove sediment sample and its application as a laundry detergent additive[J].Process Biochemistry,2006,41:1239-1243.
[9]Yinxin Z,Bo Ch,Yang Z,et al.Polar microorganisms,a potential source for new natural medicines:a review[J].Acta Microbiologica Sinica,2008,48(5):659-700.
[10]林念煒,張銳,趙晶,等.南極產(chǎn)低溫蛋白酶菌株Marinobacter sp.R2的發(fā)酵條件及酶學(xué)性質(zhì)研究[J].廈門大學(xué)學(xué)報(bào),2004,43(6):866-869.
[11]劉靜,閔行,章驥,等.一株產(chǎn)低溫蛋白酶菌株的篩選及純酶研究[J].浙江大學(xué)學(xué)報(bào),2006,32(3):251-256.
[12]吳虹,鄭穗平.低溫微生物適應(yīng)低溫的分子機(jī)制[J].生命的化學(xué),2001,21(2):163-164.
[13]Buchanan R E,Gibbens N E.伯杰細(xì)菌鑒定手冊(cè)[M].8版.北京:科學(xué)出版社,1984:274-325.
[14]東秀珠,蔡妙英.常見細(xì)菌系統(tǒng)鑒定手冊(cè)[M].北京:科學(xué)出版社,2001:128-190.
[15]史勁松,吳奇凡,許正宏,等.中國冰川1號(hào)產(chǎn)適冷蛋白酶耐冷菌的分離鑒定及產(chǎn)酶條件[J].微生物學(xué)報(bào),2005,45(2):258-262.
[16]徐國英,林學(xué)政,王能飛,等.產(chǎn)低溫蛋白酶極低菌株的篩選及Pseudoalterom onas sp.QI-1產(chǎn)蛋白酶粗酶性質(zhì)[J].生物加工過程,2010,8(2):56-60.
ABSTRACTAstrain producing the cold-adapted protease has been isolated from the frozen soil of the Tianshan Mountains,China.Based on the morphological,physiological,biochemical properties and 16S rDNA sequence analysis,the strain was identified as Pseudomonas sp.N-8.Fermentation conditions were optimized by single factor and orthogonal experiments.The optimal medium components were determined as follows:10 g/L glucose,8 g/L yeast powder,20 g/L casein,0.4 g/L KH2PO4,pH 7.0.The research on the properties of crude enzyme showed that the optimal pH and temperature for activity were 7.0 and 25℃,respectively.As to the thermal stability of the cold-adapted protease,it was stable over the range of 25~45℃.The test of pH stability indicated that the cold-adapted protease was more stable within pH 7.0~8.0.The protease activity was inhibited by Cu2+,K+,Ca2+,and Mn2+,but it was stimulated by Fe2+,Mg2+,Na+,and Zn2+.
Key wordspsychrotrophs,cold-adapted protease,enzymatic properties,Psedomonas
Screening,Identification of Pseudomonas sp.N-8 producing Cold-adapted Protease and the Study on Enzymatic Properties
Qi Dan-dan1,Yang Rui-jin2,Hua Xiao2,Shen Lian-lian1,Zhang Wen-bin2,Zhao Wei2
1(State Key Laboratory of Food Science and Technology,Jiangnan University,Wuxi 214122,China)
2(School of Food Science and Technology,Jiangnan University,Wuxi 214122,China)
碩士研究生(楊瑞金教授為通訊作者)。
*江蘇省自然科學(xué)基金“疏水納米微環(huán)境中β-半乳糖苷酶轉(zhuǎn)糖基反應(yīng)機(jī)制的研究”(編號(hào)BK2011149),以乳糖異構(gòu)化為目標(biāo)的葡萄糖異構(gòu)酶定向改造研究中高通量篩選模型的建立(其他)
2012-03-20,改回日期:2012-04-27
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
