不同土壤水分條件下欒樹光合作用的光響應
2012-09-06 04:00:36陳志成王榮榮王志偉楊吉華王華田耿兵張永濤
中國水土保持科學 2012年3期
關鍵詞:模型
陳志成,王榮榮,王志偉,楊吉華,王華田,耿兵,張永濤
(山東農業大學林學院,山東省土壤侵蝕與生態修復重點實驗室,271018,山東泰安)
不同土壤水分條件下欒樹光合作用的光響應
陳志成,王榮榮,王志偉,楊吉華,王華田,耿兵,張永濤?
(山東農業大學林學院,山東省土壤侵蝕與生態修復重點實驗室,271018,山東泰安)
采用光合作用光響應直角雙曲線模型、非直角雙曲線模型和直角雙曲線修正模型對欒樹在不同土壤水分條件下的光響應曲線進行擬合,探討不同光響應模型對欒樹的適用性及欒樹光合特性對土壤水分和光照強度的響應規律。結果表明:1)3個模型都能較好地擬合欒樹的光合光響應過程,其中直角雙曲線修正模型能擬合在光抑制條件下的光響應過程,非直角雙曲線模型和直角雙曲線修正模型要優于直角雙曲線模型;2)欒樹對土壤水分和光照強度的適宜范圍較廣,土壤相對含水量在41.5%~93.3%范圍內,凈光合速率、表觀量子效率均相對較高,光合有效輻射強度在600~2 000 μmol/(m2·s)范圍內,凈光合速率和水分利用效率均能獲得較高水平。
欒樹;土壤水分;光合作用光響應;光響應模型;蒸騰作用;水分利用效率
光和土壤水分是影響植被生長及分布的重要生態因子[1]。土壤水分虧缺會嚴重影響植物的光合作用,限制樹木的正常生長,并且強光也會抑制植物的光合作用,使光合效率下降[2]。通過光響應曲線可得出多個生理參數(包括表觀量子效率、最大凈光合速率、光補償點、光飽和點以及暗呼吸速率)[3-6],這些參數是各尺度植物生理生態學過程研究的基礎[3]。目前,國內外多采用直角雙曲線與非直角雙曲線模型對植物的光響應過程進行擬合[3];但在使用這2種光響應模型時,求得的光飽和點低于實際值,最大凈光合速率大于實際值,且無法擬合光抑制情況下的光響應曲線[4-5]。葉子飄等[5,7-8]提出了一種直角雙曲線修正模型,此模型可以克服上述2種模型的缺陷[4-5],但此模型目前主要應用于農作物和草本植物[7-8]的光合作用光響應模擬,在木本植物方面應用較少。
欒樹(Koelreuteria paniculataLaxm.)為無患子科欒樹屬喬木,喜光、耐干旱,多分布在我國北方[9]。目前關于欒樹的研究多限于苗木繁育、園林應用[10-11]等方面,涉及生理生態方面的報道較少,尤其是在不同土壤水分條件下欒樹的光合性能還不清楚。筆者采取盆栽控水的試驗方法,研究不同土壤水分條件下欒樹光合作用的光響應,并采用光響應直角雙曲線模型、非直角雙曲線模型和直角雙曲線修正模型對光響應曲線進行擬合,以探討不同光響應模型擬合欒樹在不同水分條件下光響應曲線的適用性及欒樹光合特性對土壤水分和光照強度的響應規律,為了解欒樹的光合生理生態特征及在北方山區植被恢復與重建中的應用提供理論依據。
1 試驗地概況
試驗地設在山東省泰安市東南部的山東農業大學南校區林學實驗站(E 116°08',N 36°06'),海拔154m。極端最低氣溫為-22℃,最高氣溫為40℃,無霜期186.6 d。降雨多集中在7—9月,多年平均降水量741.8mm。≥10℃年積溫2 350~4 777℃,年平均氣溫12.9℃,年均相對濕度65%,年均日照時間2 583 h。土壤類型以褐土、棕壤為主。
2 研究方法
2.1 試驗材料與方法
選擇生長一致的2年生欒樹苗木為試驗材料。2011年3月進行盆栽,8月正式試驗。花盆土壤密度為1.34 g/cm3、田間持水量30%(環刀法)。采用人為給水和自然耗水相結合的方法獲得不同土壤含水量。試驗開始前1 d澆水,使土壤含水量飽和,然后自然耗水。用質量含水量(鋁盒烘干法)與田間持水量的比值獲得土壤相對含水量(RWC/%)。共獲得了4個土壤水分梯度,土壤相對含水量分別為93.3%、73.8%、41.5%和28.5%。試驗期間搭建簡易遮雨棚防止雨水進入土壤。
選取3盆生長健壯較一致的盆栽苗(每盆1株),在晴朗或少云天氣,用CIRAS-2型便攜式光合儀于上午08:30—11:30之間測定欒樹苗木中上部成熟葉,每個葉片測定1次光響應曲線,一株樹測2片葉子。測定時大氣 CO2摩爾分數(385±10)μmol/mol,大氣溫度 25~28℃,相對濕度(60±10)%。設人工光照強度 2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、250、150、100、50、20、0 μmol/(m2·s),每個光照強度測120 s。記錄光合有效輻射強度(PAR/(μmol/(m2·s)))、凈光合速率(Pn/(μmol/(m2·s)))、蒸騰速率(Tr/(mmol/(m2·s)))等光合生理參數,水分利用效率(WUE/(μmol/mmol))利用公式WUE=Pn/Tr計算。
用Excel軟件得出凈光合速率的光響應曲線,根據曲線走勢估計光飽和點(LSP/(μmol/(m2·s)))、最 大 凈 光 合 速 率 (Pmax/(μmol/(m2·s)))[4,6],作為光響應特征曲線的實測值;對光響應曲線的初始部分(PAR<160 μmol/(m2·s))進行線性回歸,由線性方程求得表觀量子效率(Φ/(mol/mol))、光補償點(LCP/(μmol/(m2·s)))和暗呼吸速率(Rd/(μmol/(m2·s)))[4,6],作為實測值與模型擬合值進行比較分析。
2.2 模型擬合
用SPSS17.0、Excel軟件對凈光合速率光響應數據進行非線性回歸擬合。
2.2.1 直角雙曲線模型 該模型表達式[4,7]為
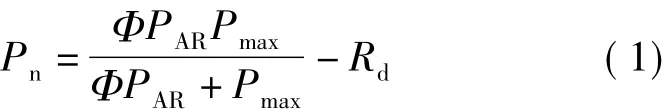
該模型光補償點計算公式為
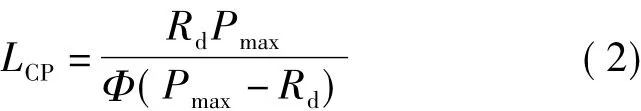
直線y=Pmax與弱光下的線性方程相交,交點對應x軸的值即為光飽和點[4,8]。
2.2.2 非直角雙曲線模型 該模型表達式[4,12]為
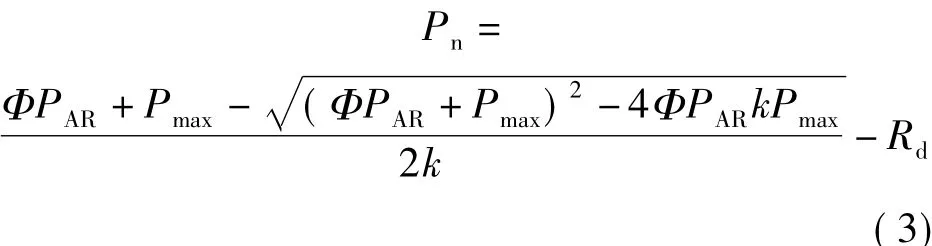
式中k為模型曲線曲角,量綱為1。
該模型光補償點計算公式為
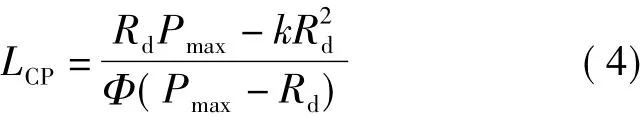
光飽和點的求算方法同式(2)。
2.2.3 直角雙曲線修正模型 該模型表達式[4-7]為
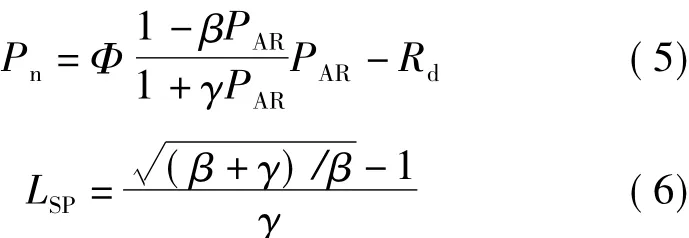
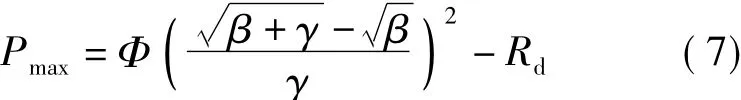
式中β、γ為模型系數。
3 結果與分析
3.1 欒樹光合作用光響應的模擬
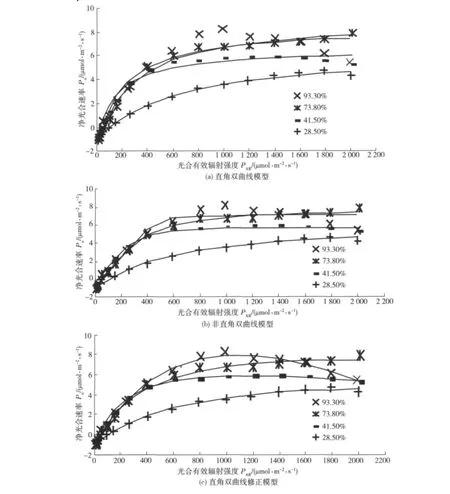
圖1 3個模型對欒樹光合速率光響應曲線的模擬Fig.1 Simulation of photosynthetic rate-light response curves of Koelreuteria paniculata Laxm.by 3models
用直角雙曲線模型、非直角雙曲線模型和直角雙曲線修正模型3個模型對欒樹光響應曲線的模擬結果見圖1,光響應過程特征參數的擬合結果見表1。可知:土壤相對含水量在28.5% ~73.8%范圍內,3個模型均能較好地模擬欒樹光合速率光響應過程,模型擬合的決定系數R2都超過0.9,其中直角雙曲線修正模型的R2在0.996以上;當土壤相對含水量為93.3%時,欒樹的光合作用在強光下發生了明顯的光抑制,此時只有直角雙曲線修正模型的擬合結果較好,且R2為0.999,優于另外2個模型。這是因為直角雙曲線和非直角雙曲線模型在理論上是一條沒有極值的漸近線,在擬合光抑制情況下的光合光響應過程會產生較大的偏差。
由表1可以看出,3個模型均能較好地擬合欒樹光合作用的表觀量子效率Φ、光補償點LCP和暗呼吸速率Rd,綜合比較Φ、LCP和Rd與實測值的接近程度,可以發現,非直角雙曲線模型、直角雙曲線修正模型要優于直角雙曲線模型,而非直角雙曲線模型與直角雙曲線修正模型之間難分優劣。2個傳統模型對最大凈光合速率Pmax的擬合值大于實測值,對光飽和點LSP的擬合值小于實測值,而直角雙曲線修正模型對Pmax、LSP的擬合結果都較好。

表1 欒樹光合光響應參數Tab.1 Data of photosynthesis-light response parameters of Koelreuteria paniculata Laxm.
3.2 欒樹葉片光合作用的光響應
圖2為欒樹凈光合速率的光響應曲線。可以看出:在較低光照強度范圍內(光合有效輻射強度PAR<600 μmol/(m2·s)),各土壤水分條件下的凈光合速率Pn對PAR的響應都反應敏感,Pn隨著PAR的增強較快上升,但是在PAR>600 μmol/(m2·s)范圍內,光響應過程因土壤水分的不同具有較大差別,即:當土壤相對含水量為93.3%時,Pn在PAR為1 000 μmol/(m2·s)左右處達到飽和,之后Pn隨PAR的增強而降低,表現出明顯的光抑制現象;土壤相對含水量為73.8%與41.5%時,在PAR>800 μmol/(m2·s)范圍內,Pn都變化緩慢;土壤相對含水量為28.5%時,隨PAR的增大,Pn一直在緩慢上升,PAR>1 800 μmol/(m2·s)之后,Pn出現下降。值得注意的是,在土壤水分較高(RWC為93.3%)時出現了最高的凈光合速率,Pn實測值為 8.25 μmol/(m2·s),之后又出現了明顯的光抑制現象。這說明在土壤含水量較高時,欒樹可能出現了水漬脅迫,但是光照強度在600 ~1 600 μmol/(m2·s)范圍內時又可以維持較高的光合生產力。在RWC為28.5%時,Pn最大值為4.7 μmol/(m2·s),同土壤水分條件下,遼東楤木(Aralia elata)Pn最大值約為 2 μmol/(m2·s)[12],核桃(Juglans regia)Pn最大值約為3 μmol/(m2·s)[13],黃櫨(Cotinus coggygriavar.cinerea)Pn最大值約為0.5 μmol/(m2·s)[14],可知,在低土壤水分條件下,欒樹光合生產力相對較高。結合表1,土壤相對含水量在41.5% ~93.3%范圍內時,凈光合速率、表觀量子效率均相對較高。欒樹對光照強度的適應范圍較廣,同一土壤水分條件下,在600~2 000 μmol/(m2·s)光照強度范圍內Pn值均較高。
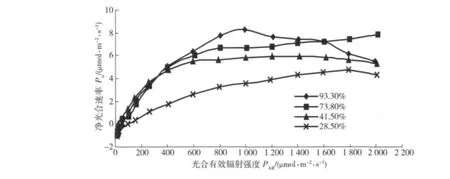
圖2 欒樹凈光合速率的光響應曲線Fig.2 Light responses of net photosynthetic rate of Koelreuteria paniculata Laxm.
表觀量子效率Φ值高,則植物在弱光下轉換利用光能的效率就高[2]。從表1可見,欒樹表觀量子效率在0.009 9~0.021 3mol/mol之間,低于一般植物的表觀量子效率(0.03 ~0.05mol/mol)[4,14]。筆者發現,土壤相對含水量為93.3%的Φ值最大,之后隨土壤水分的降低,Φ值也呈降低趨勢,說明欒樹在高土壤含水量下轉化弱光的光能效率高。
光補償點LCP越小,表明植物利用弱光的能力越強,光飽和點LSP越大,則植物利用強光的能力越大[2,15]。由表1 可知,隨著土壤含水量的降低,LCP先降低后升高,LSP一直升高,在土壤相對含水量為28.5%時達到最高。這說明在土壤含水量低時欒樹對強光的適應能力較強,這與其他一些植物在低土壤含水量時發生光抑制現象不同[4,16]。
3.3 欒樹葉片蒸騰速率的光響應
圖3為欒樹蒸騰速率的光響應曲線。可以看出,不同土壤水分條件下欒樹葉片的蒸騰速率Tr差別較大,土壤相對含水量為73.8%時的Tr最大,最大值為2.8mmol/(m2·s),其次是RWC為41.5%時,說明土壤含水量過高或過低都會導致欒樹Tr降低。土壤相對含水量為73.8%和41.5%時,Tr對光合有效輻射強度PAR的響應過程相似,即在PAR<400 μmol/(m2·s)的弱光范圍內,隨著PAR的增強,Tr上升較快,但超過此光照強度以后,Tr無明顯變化。土壤相對含水量為93.3%與28.5%時,Tr隨PAR的增強沒有明顯變化。在PAR>400 μmol/(m2·s)范圍內,各水分梯度下的Tr都無明顯變化,這說明欒樹蒸騰作用受土壤水分的影響大,受光照強度的影響小。
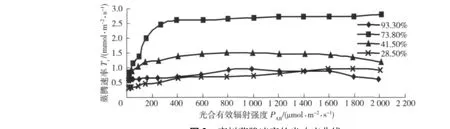
圖3 欒樹蒸騰速率的光響應曲線Fig.3 Light responses of transpiration rate of Koelreuteria paniculata Laxm.
3.4 欒樹葉片水分利用效率的光響應
圖4為欒樹水分利用效率的光響應曲線。在低光照強度PAR<400 μmol/(m2·s)范圍內,WUE對PAR的增強反應敏感,升高迅速,隨PAR繼續增大,WUE隨PAR增強的變化減弱,這說明欒樹WUE對PAR的適應范圍較廣,并在強光下可以維持較高的水分利用效率。不同土壤水分條件下欒樹葉片的WUE差別明顯:土壤相對含水量為93.3%時,WUE達到最高水平(最大值為8.86 μmol/mmol),這是在此水分梯度下Pn較高而Tr較低所致;土壤相對含水量在28.5%時,WUE高于另外2個水分梯度的WUE值,這是因為在此水分梯度下Tr較低所致。欒樹葉片在土壤水分含量最高和最低時水分利用效率反而很高,這與其他一些植物葉片水分利用效率在光響應過程中的結果不同,如遼東楤木在土壤相對含水量為56%時達到最高[12],小葉扶芳藤(Euonymus fortuneivar.radicans Sieb)在土壤相對含水量為72.3%時達到最高[15],五 葉 爬 山 虎 (ParthenocissusquinquefoliaPlanch)在土壤相對含水量為50.1%時達到最高[16]。
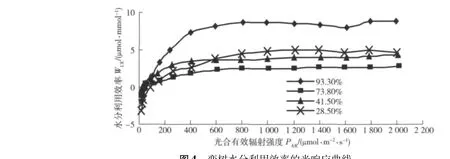
圖4 欒樹水分利用效率的光響應曲線Fig.4 Light responses of water use efficiency of Koelreuteria paniculata Laxm.
4 結論
1)直角雙曲線模型、非直角雙曲線模型和直角雙曲線修正模型都能較好地擬合欒樹的光合光響應過程,其中直角雙曲線修正模型能較好地擬合欒樹在光抑制條件下的光響應過程。綜合比較3個模型,非直角雙曲線模型與直角雙曲線修正模型要優于直角雙曲線模型。
2)在土壤含水量高時(土壤相對含水量為93.3%),欒樹出現了明顯的光抑制現象,但是此水分條件下在600~1 600 μmol/(m2·s)光照強度范圍內可以維持較高的光合生產力,并且表觀量子效率、水分利用效率都是最大;土壤含水量低時(土壤相對含水量為28.5%),欒樹光合生產力相對較高。說明欒樹既具有一定的耐濕能力,又具有一定的耐旱生產力,是對土壤水分適應性較強的植物。
3)土壤相對含水量在41.5% ~93.3%范圍內時,凈光合速率、表觀量子效率均較高,且欒樹對光照強度的適應范圍較廣,在600~2 000 μmol/(m2·s)光照強度范圍內凈光合速率、表觀量子效率均能獲得較高值。
5 參考文獻
[1]夏江寶,張光燦,劉剛,等.不同土壤水分條件下紫藤葉片生理參數的光響應[J].應用生態學報,2007,18(1):30-34
[2]韓剛,趙忠.不同土壤水分下4種沙生灌木的光合光響應特性[J].生態學報,2010,30(15):4019-4026
[3]張彌,吳家兵,關德新,等.長白山闊葉紅松林主要樹種光合作用的光響應曲線[J].應用生態學報,2006,17(9):1575-1578
[4]郎瑩,張光燦,張征坤,等.不同土壤水分下山杏光合作用光響應過程及其模擬[J].生態學報,2011,31(16):4499-4508
[5]葉子飄,于強.一個光合作用光響應新模型與傳統模型的比較[J].沈陽農業大學學報,2007,38(6):771-775
[6]許大全.光合作用效率[M].上海:上海科學技術出版社,2002:33-33
[7]葉子飄,王健林.植物光合-光響應模型的比較分析[J].井岡山學院學報:自然科學版,2009,30(4):9-13
[8]葉子飄,于強.光合作用光響應模型的比較[J].植物生態學報,2008,32(6):1356-1361
[9]任憲威.樹木學[M].北京:中國林業出版社,1997:415
[10]吉付印,邵明麗,何珍.欒樹培育技術及園林應用[J].現代園藝,2011(8):37-38
[11]潘曉鋒.欒樹林圃兩用林豐產高效培育技術[J].現代農業科技,2009(13):214-216
[12]陳建,張光燦,張淑勇,等.遼東楤木光合和蒸騰作用對光照和土壤水分的響應過程[J].應用生態學報,2008,19(6):1185-1190
[13]李小磊,張光燦,周澤福,等.黃土丘陵區不同土壤水分下核桃葉片水分利用效率的光響應[J].中國水土保持科學,2005,3(1):43-47
[14]劉剛,張光燦,劉霞.土壤干旱脅迫對黃櫨葉片光合作用的影響[J].應用生態學報,2010,21(7):1697-1701
[15]張淑勇,周澤福,夏江寶,等.不同土壤水分條件下小葉扶芳藤葉片光合作用對光的響應[J].西北植物學報,2007,27(12):2514-2521
[16]張淑勇,張光燦,陳建,等.土壤水分對五葉爬山虎光合與蒸騰作用的影響[J].中國水土保持科學,2006,4(4):62-66
Light response of photosynthesis ofKoelreuteria paniculataLaxm.under different soil water conditions
Chen Zhicheng,Wang Rongrong,Wang Zhiwei,Yang Jihua,Wang Huatian,Geng Bing,Zhang Yongtao
(Forestry College,Shandong Agricultural University,Shandong Provincial Key Laboratory of Soil Erosion and Ecological Restoration,271018,Tai'an,Shandong,China)
The light responses of photosynthesis ofKoelreuteria paniculataLaxm.were studied under 4 different soil water conditions,and the light response curves of photosynthesis were fitted and analyzed by rectangular hyperbolamodel,non-rectangular hyperbolamodel and amodifiedmodel based on rectangular hyperbolamodel.Then the bestmodel forKoelreuteria paniculataLaxm.was selected.Photosynthetic characteristic were studied under different soil water conditions and different light intensity.The results show that:1)The light response of photosynthesis were well fitted by the threemodels,and themodified rectangular hyperbolamodel could well fit light response of photosynthesis of photo-inhibition.Nonrectangular hyperbolamodel andmodified rectangular hyperbolamodel is better than rectangular hyperbolamodel.2)Koelreuteria paniculataLaxm.had strong adaptability to soil water and light conditions.Higher photosynthetic rate(Pn)and apparent quantum yield(Φ)were observed in the region where the relative soil water contents(RWC)were from 41.5%to 93.3%.HigherPnand water use efficiency(WUE)were observed in the region where the photosynthetic active radiation(PAR)were from 600 to 2 000 μmol/(m2·s).
Koelreuteria paniculataLaxm.;soil water content;light response of photosynthesis;light responsemodel;transpiration;water use efficiency
2011-12-09
2012-03-19
世界銀行貸款山東省生態造林項目“干旱瘠薄山地樹種及造林模式選擇研究”(SEAP-KY-1)
陳志成(1986—),男,碩士研究生。主要研究方向:林業生態工程。E-mail:wwchenzhicheng2006@163.com
?責任作者簡介:張永濤(1972—),男,博士,副教授,碩士生導師。主要研究方向:林業生態工程。E-mail:yongtaozhang@126.com
(責任編輯:宋如華)
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
