江西省黃鼬和鼬獾狂犬病流行病學監測*
2012-08-21 06:51:52趙敬慧張守峰苗富春王林棟李易潞扈榮良
中國人獸共患病學報 2012年11期
趙敬慧,張守峰,劉 曄,陳 奇,苗富春,2,王林棟,李易潞,扈榮良
2.吉林農業大學動物科學技術學院,長春 130118
狂犬病(Rabies)是由狂犬病病毒(Rabies virus,RABV)引起的一種古老的人獸共患傳染病,死亡率幾乎為100%。狂犬病在我國大部分地區流行,嚴重危害公共衛生安全。在我國,除犬外野生動物可能也是重要的傳播宿主和貯存宿主,野生動物狂犬病尚無系統研究。鼬獾(Melogalemoschata)是一種在我國華東、東南丘陵地區廣泛分布的小型鼬科動物。本研究室對江西、浙江、安徽等地區野生動物鼬獾進行的狂犬病流行病學調查表明,上述地區鼬獾種群內存在狂犬病的獨立傳播,與人狂犬病關系密切[1-2];并有一株鼬獾狂犬病毒變異株JX09-17(fb)與浙江地區犬源分離株同源性較高,可能與犬狂犬病溢出有關[3]。
該地區另一種數量較大夜行野生動物——黃鼬(Mustelasibirica),俗稱黃鼠狼,廣泛分布于我國各地區,生活習性與鼬獾相似,晝伏夜出,與鼬獾同為夜行動物。黃鼬狂犬病在波多黎各、格林納達、多米尼加、古巴、南非和加勒比海岸地區均有報道,而且黃鼬分離株與犬源分離株核酸同源性較高,不同地區黃鼬分離株彼此差異較大,可能是分別感染了不同的狂犬病病毒株[4-9]。國內并沒有黃鼬狂犬病的報道,為了解我國黃鼬是否攜帶狂犬病病毒,以及黃鼬、鼬獾、犬3種動物之間是否存在交叉感染,針對江西省部分地區野生動物黃鼬及鼬獾進行狂犬病流行病學監測,確定其狂犬病感染狀況,為我國狂犬病的防治策略的制定提供依據。
1 材料與方法
1.1 樣品采集2011年11月到2012年2月,在江西省共收集黃鼬腦組織樣品1 102份,鼬獾腦組織樣品210份,見表1。

表1 黃鼬、鼬獾腦組織樣品來源地區分布Tab.1 Original place distribution of samples
1.2 直接免疫熒光檢測(FAT) 按直接免疫熒光法進行狂犬病病原檢測[11]。取小腦和腦干組織少許,在載玻片上制備組織觸片;腦組織觸片自然風干,置于80%丙酮溶液內固定(4℃,20 min),取出自然風干;以PBS(0.01 mol/L,p H7.4)稀釋FITC標記的抗狂犬病病毒核蛋白單克隆抗體 (由本實驗室制備)為工作液;將觸片置于染液內,37℃染色60 min;以洗液(1L PBS中加0.5 m L Tween-20)浸洗染色后觸片,洗滌3次;洗滌后滴加80%甘油1~2滴,加蓋玻片后熒光顯微鏡下觀察。陰性對照為正常鼠腦組織,陽性對照為狂犬病病毒BD06株鼠腦毒。
1.3 狂犬病病毒分離 采用小鼠腦內接種方法[10],對FAT檢測陽性的樣品進行病毒分離。制成10%懸液,于4℃以2 000 r/min離心20 min,取上清接種。1~3日齡昆明種乳鼠(購自長春生物制品研究所實驗動物中心),約10只/窩,乳鼠和母鼠同籠飼養。乳鼠腦內接種0.03 m L/只,觀察5~21 d。取發病或死亡的小鼠腦組織用于FAT和RTPCR檢測。
1.4 N基因和G基因的擴增與序列測定 狂犬病陽性腦組織,TRIzol法提取總RNA,以特異引物進行反轉錄,用2對引物進行PCR,擴增狂犬病病毒核蛋白和糖蛋白全基因序列,反應體系為:模板2 μL,10× PCR buffer 5μL,d NTP Master Mix 4 μL,Ex-taq DNA 聚合酶0.5μL,上下游引物各1 μL,雙蒸水補至總體積50μL。反應條件為:95℃變性3 min后,于95℃30 s,52℃30 s,72℃100 s循環擴增30次,最后72℃延伸10 min。陰性對照為正常鼠腦組織,陽性對照為狂犬病病毒BD06株。PCR產物經1%瓊脂糖凝膠電泳鑒定。如擴增獲得1.7 kb(N基因)與1.8 kb(G基因)左右條帶,回收PCR產物,送Ta KaRa公司,用擴增引物對PCR產物直接測序。拼接后核蛋白基因ORF(1 353 bp)及糖蛋白基因ORF(1 575 bp),見表2。

表2 N基因和G基因擴增引物Tab.2 Primers for amplification of N and G genes
1.5 DNA序列與分析 文件轉換使用Clustal W軟件包,用MEG4軟件以鄰位相連法構建系統發生樹。本研究中用于比較分析的狂犬病毒N基因和G基因序列來源于GenBank,見圖3,各毒株信息:GenBank登錄號/毒株/宿主/來源地區/分離時間。
2 結 果
2.1 樣品檢測與病毒分離 1 102份黃鼬腦組織樣品FAT檢測結果呈陰性。210份鼬獾腦組織樣品FAT陽性4份,可見明亮的蘋果綠或黃綠色熒光顆粒,呈不同形狀和大小。陽性鼬獾樣品腦內接種3日齡乳鼠,共分離到4株狂犬病毒,分別命名為JX12-64、JX12-67、JX12-102、JX12-234,這4株街毒在乳鼠腦內接種后在7~15 d發病。發病乳鼠腦組織FAT呈陽性,見圖1。
2.2 基因擴增產物鑒定 取5μL PCR產物進行1.0%瓊脂糖凝膠電泳,結果4株狂犬病病毒在1.7 kb及1.8 kb處均可見目的條帶,與預期大小相符,見圖2。
2.3 GenBank登錄號 4株新分離鼬獾源狂犬病病毒N基因和G基因序列已錄入GenBank,登錄號(圖3):JX12-64 N基因JQ950446,G基因JQ950447;JX12-67 N 基因JQ950448,G 基因JQ950449;JX12-102 N基因JQ950450,G基因JQ950451;JX12-234 N基因JQ950452,G基因JQ950453。
2.4 基于N基因和G基因構建系統進化樹分析見圖3,JX12-64,JX12-67,JX12-102,JX12-234,這4株狂犬病病毒同屬基因1型,病毒之間N基因和G基因的同源性較高,分別為99.6%~100%和99.7%~100%,與浙江鼬獾分離株(ZJ-LA,F02等)和部分江西其他鼬獾分離株(JX08-45,JX08-48,JX08-47)同源性也較高,分別為99.6%~100%和99.7%~100%,進化關系較近,處在同一進化分支,而且與疫苗株(CTN181和CTN7)同源性較高,分別為94.6%~95.1%和92.9%~93.6%,與浙江和福建犬源分離株(ZJ-QZ,D01,FJ008,FJ009等)同源性88.2%~88.8%和87.6%~87.7%。

圖1 FAT檢測黃鼬和鼬獾腦涂片(A)鼬獾陽性腦組織涂片;(B)正常黃鼬腦組織涂片;(C)陰性對照;(D)陽性對照Fig.1 FAT of ferret badger and yellow mongoose brainA:Brain of ferret badger with suspected rabies;B:Yellow mongoose brain;C:Negative control;D:Positive control
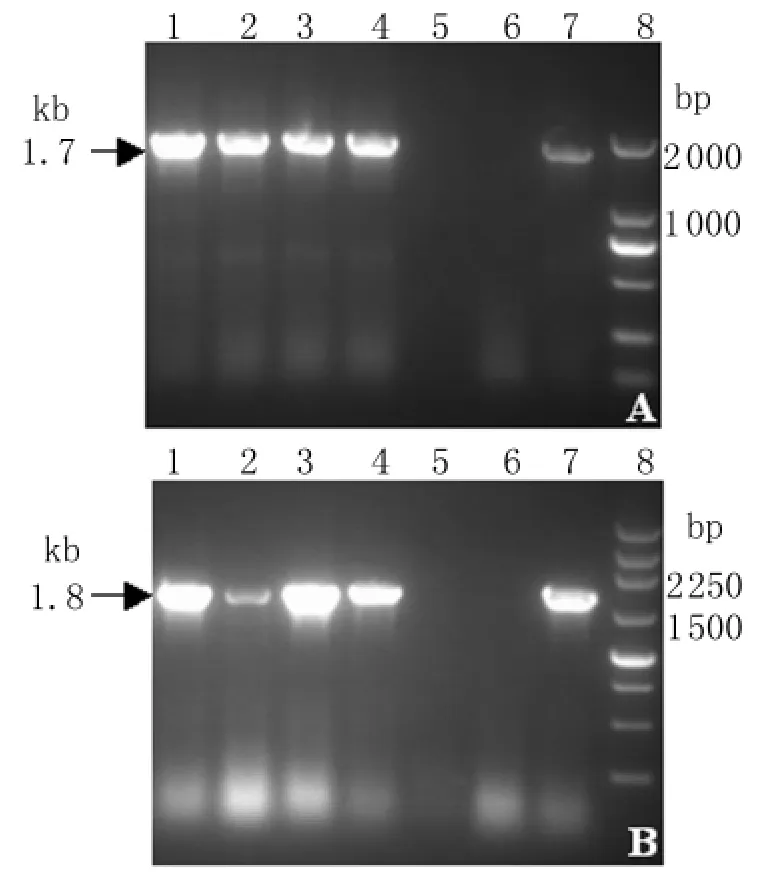
圖2 鼬獾狂犬病病毒N基因和G基因RT-PCR結果(A)N 基 因 1~4:JX12-64,JX12-67,JX12-102,JX12-234;5:空白對照;6:陰性對照;7:陽性對照;8:DL2000 Marker;(B)G 基 因 1~4:JX12-64,JX12-67,JX12-102,JX12-234;5:空白對照;6:陰性對照;7:陽性對照;8:250 bp MarkerFig.2 RT-PCR results of N and G genes from ferret badger rabies virus(A)N-gene;Lane 1 to 4:JX12-64,JX12-67,JX12-102,and JX12-234,respectively;5:PCR-negative control;6: Uninfected brain negative control;7:Positive control;8:DL2000 marker(B)G-gene.Lane 1 to 4:JX12-64,JX12-67,JX12-102,and JX12-234,respectively;5:PCR-negative control;6: Uninfected brain negative control;7:Positive control;8:250 bp marker
3 討 論
黃鼬狂犬病主要報道于加勒比海沿岸地區,最早報道于1950年波多黎各,而后在古巴、格林納達、多米尼加和南非等地也有報道[4-9]。我國并沒有黃鼬狂犬病的相關報道,本研究首次對我國江西省黃鼬和鼬獾進行了狂犬病流行病學監測,不僅對該地區黃鼬和鼬獾感染狂犬病情況有初步了解,同時,為野生動物狂犬病相關研究提供流行病學數據,也為野生動物狂犬病控制提供依據。

圖3 基于我國RV流行毒株N基因(A)和G基因(B)構建系統進化樹(A)N基因;(B)G基因;各毒株信息:GenBank登錄號/毒株/宿主/來源地區/分離時間Fig.3 The neighbor-joining phylogenetic tree of representative Chinese rabies virus isolates,using the N and G gene sequencesA:N gene;B:G gene;Information of RV:Gen-Bank no./isolate/host/origin/isolation date
黃鼬與鼬獾腦組織樣品主要來源于江西省的新建縣、安義縣、永修縣、高安縣、南昌縣、景德鎮等地區,集中在江西省北部環鄱陽湖地區。對鄱陽湖地區野生動物黃鼬和鼬獾進行狂犬病流行病學監測,一方面因為這一地區鼬獾種群內存在狂犬病的流行,黃鼬與鼬獾都是夜行動物,有交叉感染的可能;另一方面鄱陽湖近年來持續干旱,枯水期提前,豐水期推遲,2011年年初到年底,湖區水域面積減少近三分之二,甚至部分地區出現飲水困難的情況,生態環境的變化可能導致野生動物的活動范圍與區域的改變,也可能會增加狂犬病擴散傳播的幾率。本次監測結果顯示,1 102份黃鼬樣品,FAT結果都為陰性,210份鼬獾樣品,FAT陽性4份,經MIT分離獲得4株狂犬病病毒。對這4株病毒N基因和G基因遺傳進化分析,新分離4株鼬獾源狂犬病病毒同屬基因1型,同源性較高,與浙江省鼬獾分離株(ZJLA,F02等)和江西省其它鼬獾分離株(JX08-45,JX08-48,JX08-47)同源性在96%以上,遺傳進化關系也較近,處在同一進化分支,但與浙江和福建犬源分離株(ZJ-QZ,D01,FJ008,FJ009等)同源性88%左右,與疫苗株CTN181和CTN7同源性較高。CTN181是上世紀50年代分離自我國山東淄博的疫苗株,與廣東分離株(GN07)和貴州分離株(Guizhou_A101和 Guizhou_A103)同源性較高,遺傳進化關系也較近,而且與本研究分離的鼬獾源狂犬病病毒同處一個進化分支。這一分支的病毒曾經于上世紀60年代左右在我國廣泛流行[11],為何近年來在鼬獾種群內流行,是由犬傳播給鼬獾,還是由于鼬獾就是作為重要的貯存宿主,將病毒傳播給犬和人,這需要長期的監測分析與研究。此前有研究顯示,鼬獾源狂犬病病毒JX09-17(fb)株與江西、浙江、河北等地犬源分離株同源性較高,而與浙江和江西鼬獾分離株同源性相對較低,可能是由于犬狂犬病的溢出導致鼬獾感染[3],這仍需進一步研究來確定。
以上結果表明,采樣地區黃鼬樣品中未發現狂犬病病毒感染,而鼬獾感染狂犬病病毒陽性率較高,說明在鼬獾種群內存在狂犬病的流行。從犬與鼬獾狂犬病病毒株同源性較低的情況來看,二者交叉傳播或溢出的可能較低,黃鼬和鼬獾盡管都屬于夜行動物,但二者交互感染的可能性很低。江西省野生動物種類繁多,數量較大,因此應加強該地區野生動物狂犬病的監測。
[1]Liu Y,Zhang SF,Wu XF,et al.Ferret badger rabies origin and its revisited importance as potential source of rabies transmission in Southeast China[J].BMC Infect Dis,2010,10:234.DOI:10.1186/1471-2334-10-234
[2]Zhang SF,Tang Q,Wu XF,et al.Rabies in ferret badgers:Southeastern China[J].Emerg Infect Dis,2009,15(6):946-949.DOI:10.3201/eid1506.081485
[3]Zhang SF,Zhao JH,Liu Y,et al.Characterization of a rabies virus isolate from a ferret badger(Melogalemoschata)with unique molecular differences in glycoprotein antigenic site III[J].Virus Res,2010,149(2):143-151.DOI:10.1016/j.virusres.2010.01.010
[5]Everard CO,Everard JD.Mongoose rabies[J].Clin Infect Dis[J].1988,10(Suppl 4):S610-S614.DOI:10.1093/clinids/10.Supplement_4.S610
[6]Nel LH,Sabeta CT,von Teichman BV,et al.Mongoose rabies in southern Africa:a re-evaluation based on molecular epidemiology[J].Virus Res,2005,109(2):165-173.DOI:10.1016/j.virusres.2004.12.003
[7]Everard CO,Everard JD.Mongoose rabies in the Caribbean[J].Ann N Y Acad Sci,1992,653:356-366.DOI:10.1111/j.1749-6632.1992.tb19662
[8]Davis PL,Rambaut A,Bourhy H,et al.The evolutionary dynamics of canid and mongoose rabies virus in southern Africa[J].Arch Virol,2007,152(7):1251-1258.DOI:10.1007/s00705-007-0962-9
[9]Van Zyl N,Markotter W,Nel LH.Evolutionary history of African mongoose rabies[J].Virus Res,2010,150(1-2):93-102.DOI:10.1016/j.virusres.2010.02.018
[10]Meslin FX,Kaplan MM,Koprowski H.Laboratory techniques in rabies[M].4th ed.Geneva:World Health Organization,1996,69-80.
[11]Yu J,Li H,Tang Q,et al.The spatial and temporal dynamics of rabies in China[J].PloS Negl Trop Dis,2012,6(5):e1640.DOI:10.1371/journal.pntd.0001640
