蔥蠅對寄主植物的選擇性研究
2012-08-10 02:22:24張云霞蘇茂文趙海鵬馬曉丹
中國蔬菜 2012年4期
張云霞 王 鉦 蘇茂文 趙海鵬 馬曉丹 薛 明*
(1山東農業大學植物保護學院,山東泰安 271018;2青島出入境檢疫檢驗局,山東青島 266000)
蔥蠅〔Delia antique(Meigen)〕又稱蒜蛆,屬雙翅目花蠅科,廣泛分布于北半球溫帶地區,是世界性害蟲(Poprawski et al.,1985)。蔥蠅為寡食性害蟲,為害洋蔥(AlliuMcepaL.)、大蔥(AlliuMfistulosumL. var.giganteumMakino)、大蒜(AlliuMsativuML.)和韭菜(AlliuMtuberosumRottl. ex Spr.)等百合科蔬菜,是重要的地下害蟲,在我國以大蒜受害最重。該蟲以幼蟲蛀食寄主植物的地下部,造成地下部腐爛,整株地上部枯死;春、秋季均可為害,以春季為害最重,田間蟲株率一般可達20%~30%,嚴重時達50%以上,是目前危害我國大蒜生產的重要害蟲(王永衛和徐繼明,1990)。
對蔥蠅的研究,國外在生物學和生態學上研究較多(Ishikawa & Tsukada,1987;Ishikawa et al.,2000;Nomura & Ishikawa,2000,2001),國內在防治方面有一些報道(金運蘭 等,2007;王玉桂,2010)。蔥蠅為寡食性害蟲,其發生程度與寄主植物有密切的關系,昆蟲的產卵和取食選擇是決定其發生程度的重要因素(劉廷明 等,1989)。本試驗研究了蔥蠅在 4種不同寄主植物上發生為害情況的差異,以明確成蟲產卵選擇和幼蟲取食選擇在其中所起的作用,為掌握該蟲的田間數量動態和預測預報、加強田間蔥蠅的針對性防治提供理論依據。
1 材料與方法
1.1 蔥蠅的飼養
蔥蠅于2008年5月采自山東省泰安市郊區范鎮大蒜田,養蟲室內用大蒜連續飼養多代。溫度(20±2)℃,相對濕度60%~70%,光周期L/D=16 h/8 h。
將羽化出的成蟲分別放入養蟲籠內,籠內懸掛分別裝有牛奶、10%糖水和清水的飼養盒。在養蟲籠底部放入 1個裝有濕沙的產卵盤,并插入新鮮大蒜瓣,以誘集成蟲在沙中產卵。每天將產卵盤內的細沙倒入小盆中,倒入清水,輕輕攪拌后卵漂浮到水面上,將卵撈出后得到發育一致的卵。分批連續飼養,獲得供試驗用的發育一致的成蟲和1齡、3齡幼蟲。
試驗選用的 4種寄主植物均為山東地區普遍種植的常規品種,韭菜品種為壽光獨根紅;大蔥品種為章丘大蔥;大蒜品種為金鄉紅皮蒜;洋蔥品種為淄博紅皮洋蔥。
1.2 蔥蠅對寄主植物的選擇
1.2.1 發生情況 2010年4月上旬,在蔥蠅的發生期,在山東農業大學試驗地中選取條件一致的試驗小區4塊,每小區20 m2。每塊地十字型分為4個區域,分別移栽韭菜和苗期的洋蔥、大蔥、大蒜,罩尼龍網。然后每罩籠內接入 200頭蔥蠅成蟲(雌∶雄=2∶1)。在罩籠內上方中間懸掛盛有脫脂棉的小盤,里面分別加牛奶和清水飼喂,注意及時補充、更換。待蔥蠅幼蟲發育至2~3齡時(接成蟲后15 d),檢查不同寄主植物地下根莖部幼蟲的數量,計算蟲口數量和所占比例。
1.2.2 成蟲對寄主植物的產卵選擇 采用紗網籠罩盆栽試驗。分別在花盆(直徑9 cm,高9 cm)中移栽韭菜、苗期的洋蔥、大蔥和大蒜4種寄主植物。洋蔥、大蔥和大蒜每盆4株,韭菜每盆8株,植株冠部大小和高度基本一致,土壤表層鋪沙。在每個紗籠(60 cm×60 cm×40 cm)的4個角隅處分別放置種植4種寄主植物的花盆各3盆。然后每紗籠放入30頭己經交配且卵巢已發育成熟的雌成蟲,供其自由選擇。注意減少光線不勻的影響,試驗條件同上。6 d后采用漂浮法從不同寄主植物帶卵的細沙土中漂浮出卵和卵殼,計數。養蟲室溫度(20±2)℃,相對濕度60%~70%,光周期L/D=16 h/8 h。
1.2.3 幼蟲對寄主植物的取食選擇 取韭菜和苗期的洋蔥、大蔥、大蒜4種寄主的地下莖,洗凈,擦干,切成2 cm長的小塊,兩兩相對呈十字形放置在鋪有濾紙的玻璃培養皿(直徑20 cm)的周圍。各挑取孵化后第2天的蔥蠅1齡幼蟲100頭,2日齡的3齡幼蟲60頭,放在培養皿中間,用黑棉布覆蓋,每處理6次重復。接蟲24 h后,檢查不同寄主上幼蟲的數量,計算選擇率。

2 結果與分析
2.1 蔥蠅在4種寄主植物上的發生數量
在不同寄主植物上蔥蠅幼蟲的數量,以洋蔥最多,大蒜和大蔥次之,韭菜上最少,分別占總蟲數的40.0%、29.0%、27.0%和4.0%(圖1)。
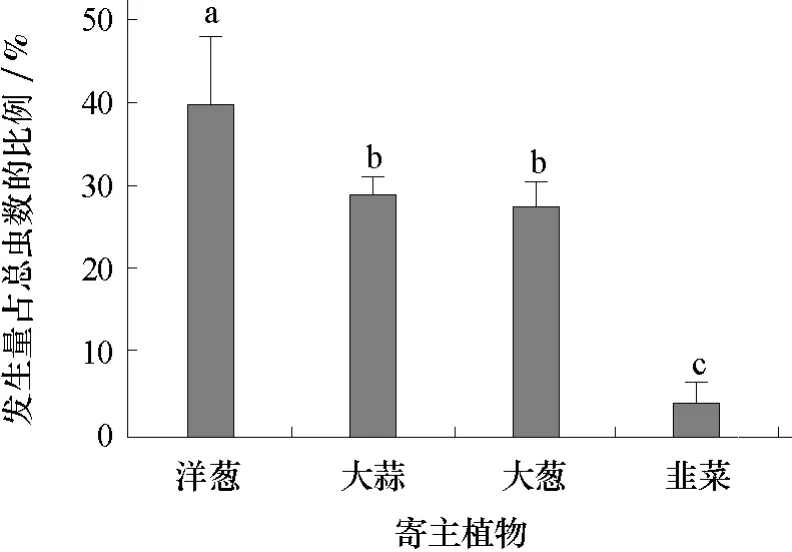
圖1 蔥蠅在4種寄主植物上的發生數量
2.2 蔥蠅成蟲對寄主植物的產卵選擇
由圖2可見,不同寄主上的落卵量依次是洋蔥(1 297.3粒)>大蒜(936.0粒)>大蔥(901.5粒)>韭菜(309.6粒),分別占總卵量的37.6%、27.2%、26.2%和9.0%。在洋蔥上的落卵量最多,韭菜上最少,大蒜和大蔥居中,差異不顯著。
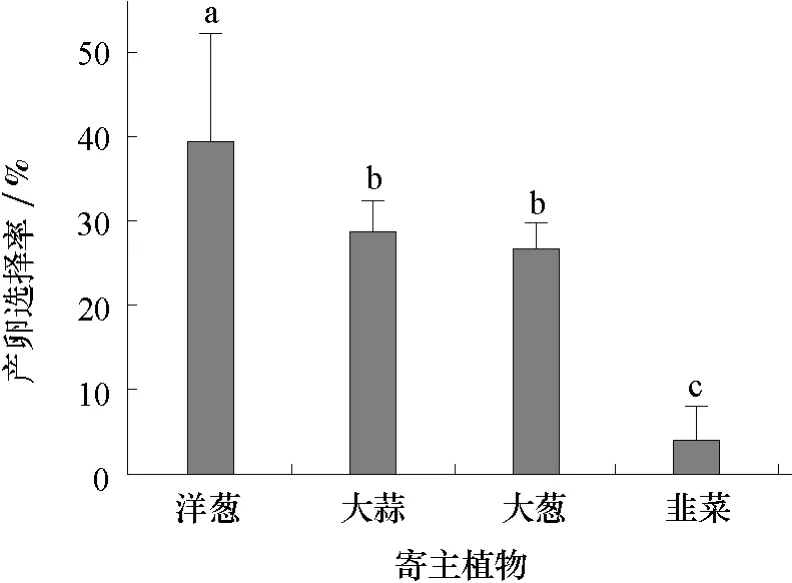
圖2 蔥蠅成蟲對4種寄主植物的產卵選擇
2.3 蔥蠅幼蟲對寄主植物的取食選擇
2.3.1 蔥蠅1齡幼蟲對寄主植物的取食選擇 由圖3可見,蔥蠅1齡幼蟲對4種寄主植物的選擇順序依次為洋蔥>大蔥>韭菜>大蒜,取食選擇率分別為76.4%、16.8%、4.2%和2.6%,其中大蒜和韭菜的差異不顯著。
2.3.2 蔥蠅3齡幼蟲對寄主植物的取食選擇 由圖4可見,接蟲24 h后不同寄主植物上的3齡幼蟲的取食選擇率依次為洋蔥(62.1%)>大蔥(23.3%)>韭菜(8.8%)>大蒜(5.8%),其中大蒜和韭菜的差異不顯著。和1齡幼蟲的取食選擇相比較,隨著蟲齡的增大,蔥蠅幼蟲對不太喜食的寄主的適應能力有一定的增強。
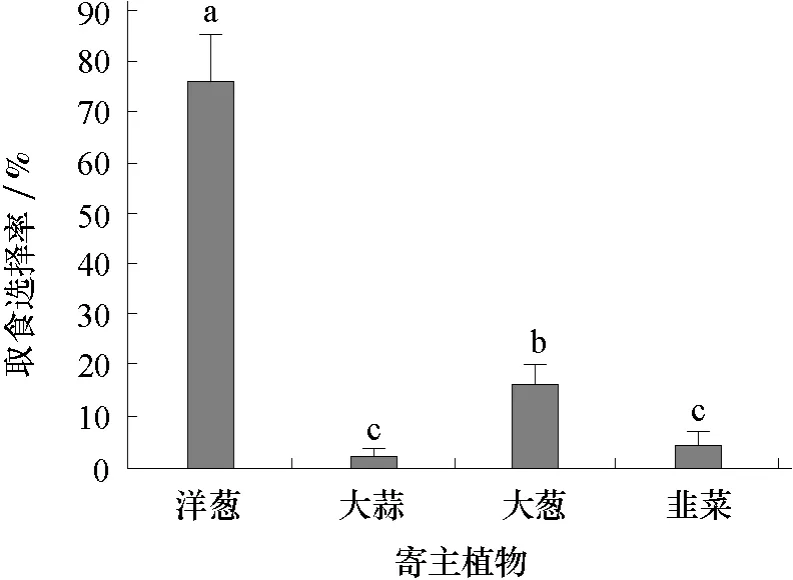
圖3 蔥蠅1齡幼蟲對4種寄主植物的取食選擇

圖4 蔥蠅3齡幼蟲對4種寄主植物的取食選擇
3 結論與討論
蔥蠅雖然為寡食性害蟲,但在田間不同種類的百合科寄主上的發生程度存在著明顯差異。本試驗結果表明,在田間罩籠的 4種寄主植物中,以洋蔥上蔥蠅的發生數量最多,其次為大蒜和大蔥,韭菜上數量最少;且盆栽蔥蠅成蟲產卵選擇試驗結果與田間罩籠試驗結果一致,說明成蟲的產卵選擇在決定蔥蠅在不同寄主的發生數量上起主要的作用。蔥蠅成蟲的嗅覺系統很發達,(Honda & Ishikawa,1987)電生理實驗證明,蔥蠅觸角嗅覺感受器對二丙二硫醚化合物有反應。據報道洋蔥散發的氣味中二丙基二硫醚及其相應的烷基硫化物等物質對蔥蠅成蟲具有引誘力并刺激其產卵(Vemon,1981);大蒜中也含有揮發性物質二烯丙基二硫及其氧化物,還有少量的二烯丙基三硫和二烯丙基四硫(周江菊,2010)。表明百合科中不同的寄主植物對蔥蠅成蟲選擇產卵的引誘力大小不同,可能與含硫化合物的組分和含量不同有關。另外蔥蠅對寄主植物的選擇除嗅覺外,視覺也起著一定的作用(Ishikawa et al.,1985;Judd & Borden,1991)。
蔥蠅幼蟲為根蛆類,相對其他昆蟲來講,活動和感覺器官簡單,僅能在小范圍內選擇取食。Soni和Finch(1979)試驗結果表明,幼蟲對寄主的選擇可能利用醇、醛、酸、酯而非單純利用硫化物作為尋找寄主的線索。所以成蟲和幼蟲在對寄主的選擇喜好上可能存在著差異。而成蟲的產卵選擇在決定對寄主植物的危害中起著更重要的作用,幼蟲的選擇僅在同種寄主植株間的選擇中起作用。蔥蠅成蟲和幼蟲均更喜歡在有破傷的鱗莖上產卵取食為害(王玉桂,2010)。
蔥蠅成蟲羽化后需大量取食來補充營養以完成生殖的需求,且成蟲壽命長,活動力強(王永衛和徐繼明,1990),這也為田間誘殺成蟲提供了良好的條件。蔥蠅是為害大蒜的重要害蟲之一,目前生產上主要在大蒜種植期采用藥劑處理土壤和幼蟲發生期田間灌根來防治蔥蠅(陳鳳春,2008),而長期大量使用化學農藥導致害蟲抗藥性增強和產品的農藥殘留(Eckenrode &Nyrop,1995),因此利用成蟲誘殺技術防治蔥蠅是實現無公害治理的重要措施之一。
陳鳳春.2008.無公害大蒜病蟲害的綜合防治.綠色植保,3(1):28-29.
金運蘭,汪彩云,史慧英.2007.蒜蛆的綜合防治.安徽農學通報,(11):160.
劉廷明,劉文英,黃厚英.1989.蔥蠅發生規律及防治技術的研究.山東農業科學,(4):15-20.
王永衛,徐繼明.1990.蔥蠅的發生及防治.新疆農墾技術,(3):17-18.
王玉桂.2010.蔥地種蠅的危害及無公害防治技術.吉林蔬菜,(5):64-65.
周江菊.2010.頂空固相微萃取氣質聯用分析大蒜揮發性風味成分.中國調味品,(9)35:95-99.
Eckenrode C,Nyrop J.1995.Onion maggot managemant in New York,Mchigan and Wisconsin.New York’s Food and Life Sciences Bulletin,144:41-42.
Honda I,Ishikawa Y.1987.Ultrasturcture of the larval cephalic sensory organs of the onion and seed-corn flies,Hylemya antiquaMeigen andH.platuraMeigen(Diptera:Anthomyiidae).Applied Entomology and Zoology,22(3):325-334.
Ishikaws Y,Tanaka S,Matsumoto Y.1985.Color preference of the onion fly with reference to ultraviolet reflection.Applied Entomology and Zoology,20(1):20-26.
Ishikawa Y,Tsukada S,Matsumoto Y.1987.Effect of temperature and photoperiod on the larval development and diapause induction in the onion fl y,Hylemya antiquaMeigen(Diptera:Anthomyiidae).Applied Entomology and Zoology,22:610-616.
Ishikawa Y,Yamashita T,Nomura M.2000.Characteristics of summer diapause in the onion maggot,Delia antiqua(Diptera:anthomyiidae).Insect Physiology,46:161-164.
Judd G J R,Borden J H.1991.Sensory interaction during trap-finding by female onion flies:imp lications for ovipositional host-p lant finding.Entomologia Experimentalis et Applicata,58:239-249.
Nomura M,Ishikawa Y.2000.Biphasic effect of low temperature on completion of winter diapause in the onion maggot,Delia antique.Insect Physiology,46:373-377.
Nomura M,Ishikawa Y.2001.DynaMic changes in cold hardimess,high-temperature tolerance and trehalose content in the onion maggot,Delia antiqua(Diptera:anthomyiidae),associated with the summer and winter diapause.Applied Entomology and Zoology,36:443-449.Poprawski T V,Robert P H,Maniania N K.1985.Susceptibility of the onion maggot to the mycotoxin destructive.Applied Entomology and Zoology,20(7):801-802.
Soni S K,Finch S.1979.Laboratory evaluation of sulphur-bearing chemicals as attractants for larvae of the onion fly,Delia antiqua(Meigen)(Diptera:Anthomyiidae).Bulletin of Entomological Research,69:291-298.
Vemon.1981.Candidate EHV technologies for the post-1980s.Vehicular Technology Conference,4:49-60.
