番茄抗病基因Ty-1的CAPS標記及檢測
2012-05-22 03:55:24李海濤杜玉麗張子君鄒慶道
中國蔬菜 2012年14期
李海濤 杜玉麗 張子君 曲 彤 鄒慶道
(1遼寧省農業科學院蔬菜研究所,遼寧 沈陽 110161;2沈陽農業大學園藝學院,遼寧 沈陽 110161)
番茄(Lycopersicon esculentum Mill.)是一種世界性經濟作物,品種多,產量高,營養豐富,用途廣泛。限制番茄生產發展和產量的主要原因在于病害的流行為害和逆境條件的制約(葉青靜 等,2009)。病毒病是番茄生產中最常見、發病最普遍的病害之一,其中的番茄黃化曲葉病毒病(Tomato yellow leaf curl virus disease,TYLCVD)是近年發生的一種暴發性、毀滅性病害,一旦發病,很難控制。番茄黃化曲葉病毒(Tomato yellow leaf curl virus,TYLCV)于1939~1940年在以色列首次發現(Pico et al.,1996),是一類由煙粉虱介導傳播的雙生病毒(geminiviruses),該病毒為雙生病毒科(Geminniviridae)菜豆金色花葉病毒屬(Begonurvints)成員(周雪平 等,2003)。番茄生長發育早期感染 TYLCV,其癥狀主要表現為植株生長緩慢或停滯,節間縮短,植株矮化,上部葉片變小、變厚、有褶皺、向上卷曲、變形、葉片邊緣至葉脈區域黃化,無法正常開花結果,下部葉片癥狀不明顯;后期感染TYLCV,染病植株僅上部葉片和新芽表現癥狀,坐果困難,果實僵化不膨大,或膨大速度極慢,果實變小,畸形果多,成熟期果實不能正常轉色、失去商品價值,導致減產或絕收。目前在中東地區、非洲、亞洲、歐洲、美洲等世界各地都有發生并且已給美國、以色列、埃及、澳大利亞等國的番茄生產造成嚴重損失,我國廣東、廣西、臺灣、上海、江蘇等地都有發生的報道(何自福 等,2007;趙統敏 等,2007),并呈現由南向北迅速蔓延的趨勢,已經成為影響番茄生產的主要限制因素之一。
目前番茄黃化曲葉病毒病的抗病基因有 Ty-1、Ty-2、Ty-3、Ty-3a、Ty-4、TY-5等,研究較多的是Ty-1、Ty-2、Ty-3基因。Zamir等(1994)認為Ty-1基因為主效基因,該基因為不完全顯性單基因,并將其定位在番茄第6號染色體的RFLP標記TG297(4 cM)和TG97(8.6 cM)之間,圖距6~10 cM。番茄黃化曲葉病毒病屬檢疫性病害,利用常規方法進行抗病育種有較大困難,利用分子標記結合常規方法能高效、準確地進行抗病材料的篩選與鑒定。本試驗旨在尋找與番茄黃化曲葉病毒病抗病基因Ty-1緊密連鎖的分子標記,并利用該標記進行抗病材料的檢測,為分子標記輔助育種奠定基礎。
1 材料與方法
1.1 材料
本試驗共用番茄材料63份。其中含抗病基因Ty-1的純合材料3份,編號為CK1、CK2、CK3,基因型為Ty-1/Ty-1,來自亞蔬中心(AVRDC);不含抗病基因Ty-1的感病純合材料3份,編號為CK4(L402母本)、CK5(L402父本)、CK6(金棚1號母本),基因型為ty-1/ty-1,來自遼寧省農業科學院蔬菜研究所番茄課題組。
抗病的F18份,編號為7~14,來自上海種都種業科技有限公司,在2010年召開的“全國抗番茄黃化曲葉病毒病品種展示會”上表現為抗病。7號為迪達,8號為迪瑞,9號為TY4220,10號為迪維斯,11號為TY4378,12號為TY4430,13號為TY1415,14號為TY3175;未知抗病性的F125份,編號為 15~39,來自國外種子公司,具體名稱不祥。未知抗病性的自交系材料24份,編號為40~63,來自遼寧省農業科學院蔬菜研究所番茄課題組。
1.2 方法
1.2.1 引物設計 番茄抗黃化曲葉病毒病抗病基因Ty-1的CAPS標記引物參照Zamir等(1994)的方法設計。引物由北京賽百盛生物技術公司合成。材料的特異性引物序列及其擴增片斷見表1。
1.2.2 PCR擴增及酶切體系 DNA提取參照Williamson等(1994)的方法,由CTAB法提取。用于PCR反應和酶切的藥品均購自tiangen生物工程公司。PCR反應總體積為20 μL,包括:模板DNA 20 ng,dNTP 0.4 μL,引物(10 U)各2.0 μL,Taq DNA聚合酶(2.5 U·μL-1)0.5 μL,Buffer 2.0μL,Mg2+2.0μL,終體積用超純水加至20 μL。PCR的反應程序為:94 ℃預變性5 min;然后94 ℃變性1 min,64.6 ℃退火1 min,72 ℃延伸1 min,35個循環;最后72 ℃延伸10 min,擴增產物4 ℃保存。酶切體系為擴增產物7 μL加入10 U的Rsa I 酶1 μL,Buffer 2 μL,終體積用超純水加至20 μL。37 ℃保溫1.5 h。PCR產物及酶切產物于2.0%瓊脂糖膠在電壓5 V·cm-1條件下電泳30 min,終結果用Goodview 染色,Bio-RAD凝膠成像系統拍照。

表1 引物序列
1.2.3 CAPS標記的獲得及抗病基因檢測 利用設計的引物對3份抗病純合材料CK1、CK2、CK3及3份感病純合材料CK4、CK5、CK6進行PCR擴增,然后將擴增產物用Rsa I酶切,獲得與抗病基因緊密連鎖的CAPS標記;利用該標記分別對F1、自交系進行抗病基因的檢測。
2 結果與分析
2.1 與Ty-1基因緊密連鎖的CAPS標記片段的獲得
6份材料的PCR擴增圖譜如圖1,酶切圖譜如圖2。
如圖1顯示,利用特異引物對6份抗病、感病材料進行PCR擴增,無論抗病材料還是感病材料均擴增出400 bp左右的片斷,經RsaⅠ酶切后(圖2),3份感病材料產生350 bp和50 bp左右的片段(但較小的50 bp的特異片段難以觀察到,可能是目標條帶產物相對較少,經酶切后小片段濃度較低所致)。3份抗病材料不能被 RsaⅠ酶切,仍呈現原來的 400 bp左右的片段。400 bp和350 bp兩個片段能將抗病材料和感病材料加以區分,是與抗病基因連鎖的標記。
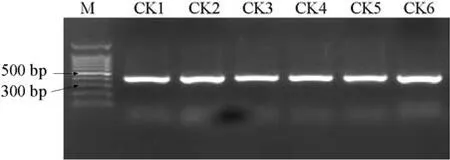
圖1 6份材料的PCR擴增圖譜
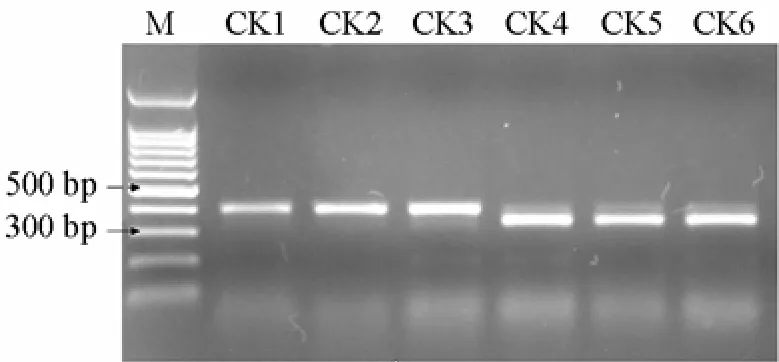
圖2 PCR擴增產物的RsaⅠ酶切圖譜
2.2 利用 CAPS標記對 8份抗病番茄 F1進行抗病基因檢測
8份抗病材料的酶切圖譜如圖3。
如圖3所示,8份抗病材料中,迪維斯、TY1415、TY31753份材料酶切后產生400、350 bp和50 bp左右的片段,說明這3份材料為含有抗病基因 Ty-1的雜交種;其他材料經酶切后只產生350 bp和50 bp左右的片段,和感病對照的擴增片段相同,說明這些材料不含抗病基因Ty-1。由于這些材料在田間均表現為抗病,說明這些材料可能含有除Ty-1以外的其他抗病基因。

圖3 8份抗病番茄F1的RsaⅠ酶切圖譜
2.3 利用CAPS標記對25份未知抗病性的番茄F1進行抗病基因檢測
25份材料酶切圖譜如圖4。
如圖4所示,25份材料中有17份材料表現為雜合抗病型,酶切后產生400、350 bp和50 bp左右的片段,應是含有抗病基因Ty-1的抗病品種;8份材料表現為純合感病基因型,酶切后產生350 bp和50 bp左右的片段,說明不含抗病基因Ty-1。
2.4 利用CAPS標記對24份番茄自交系進行抗病基因檢測
24份材料酶切圖譜如圖5所示:24份材料均存在RsaⅠ酶切位點且酶切后均產生350 bp和50 bp左右的片段,與感病對照的擴增片段相同,說明這些自交系均不含抗病基因Ty-1。
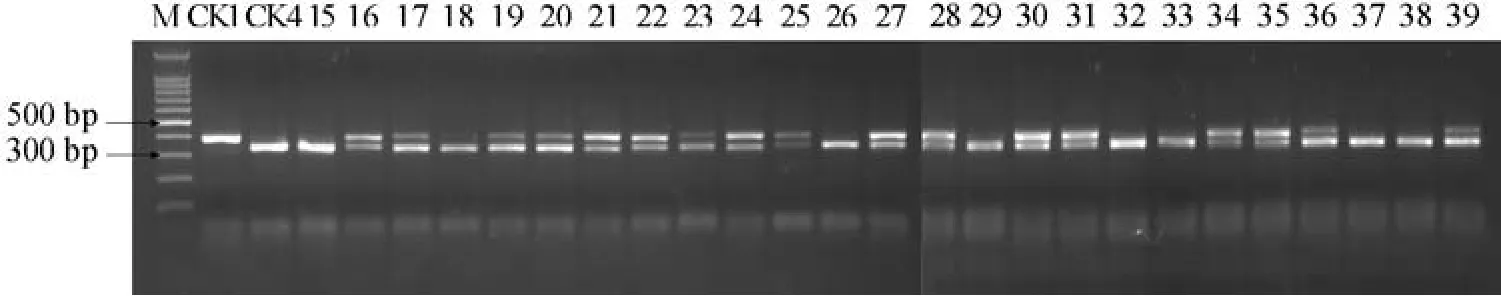
圖4 25份番茄F1的RsaⅠ酶切圖譜
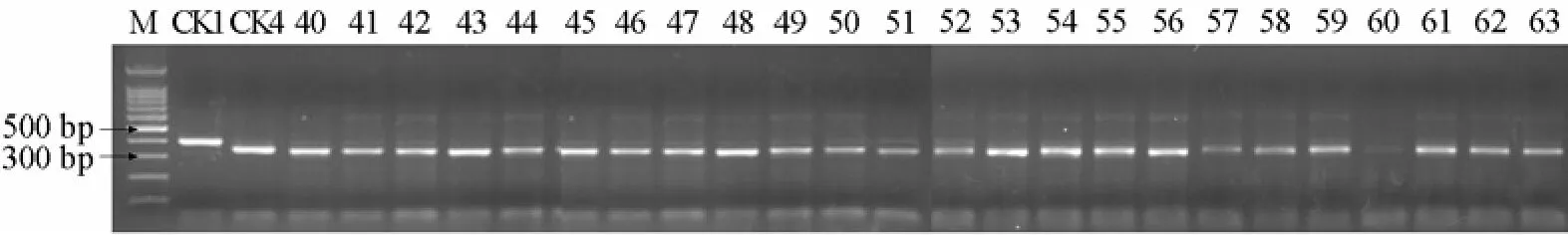
圖5 24份番茄自交系的RsaⅠ酶切圖譜
3 討論
CAPS標記作為一種特異PCR標記,不僅比RFLP、AFLP等標記快速、簡便,并且不用使用放射性同位素,減少了對人體的傷害,而且比RAPD、ISSR等PCR標記更穩定,特異性更強(于力 等,2008)。隨著分子技術的迅速發展,利用分子標記輔助育種已極大地加快了育種的速度。
本試驗找到了一個有效可用的 CAPS標記,為番茄黃化曲葉病毒病分子輔助育種奠定了基礎,但該標記與抗病基因連鎖距離的遠近,尚需田間試驗進一步驗證。利用獲得的標記共對33份番茄F1進行抗病基因的檢測,有20份材料含有抗病基因Ty-1,13份材料不含Ty-1基因,含有 Ty-1基因的抗病品種多來自國外,說明國外抗病材料比較豐富。對 24份國內骨干育種材料進行檢測,均不含抗病基因Ty-1,說明國內進行番茄抗TYLCVD育種不能直接利用以往的骨干自交系,需要進行材料創新。
不同地區的番茄黃化曲葉病毒的變異較大,導致同一個抗性材料在不同地區有不同抗性表現,甚至在同一地區不同年份間抗性表現不同。實踐表明,選育抗病品種是防治TYLCV最為經濟環保有效的方法。經過育種家多年的努力,國外在抗番茄黃化曲葉病毒病育種方面取得了顯著成就。但是目前,抗TYLCVD的商業雜交種中導入單個抗性基因的居多,當病害大規模發生時,它們的抵抗能力依然下降。提高抗性水平的策略之一就是將多個抗性基因聚合到一個品種中,而有關專家已經證明了通過聚合不同抗源的基因可以提高番茄對TYLCV的抗性(Vdavski et al.,2008)。付蓉蓉等(2011)的研究已表明,同時含有純合Ty-1和Ty-3抗性基因的番茄材料有更高更穩定的抗性。分子標記輔助選擇的最大優越性表現在抗性基因累加系的構建中,將抗性基因累加到栽培品種是育種上培育持久抗性品種的有效手段之一(余文貴 等,2009)。并且隨著番茄黃化曲葉病毒病的抗性基因 Ty-1、Ty-2、Ty-3、Ty-4及 Ty-5的定位及分子標記工作的進行(Zamir et al.,1994;Hanson et al.,2006;Ji et al.,2007,2009;Anbinder et al.,2009),育種學家可以利用相關的分子標記更加準確快速地篩選抗源材料,并結合傳統育種將這些抗性基因聚合到一個品種中,從而培育出具有更高、更廣、更持久抗性的番茄新品種(付蓉蓉 等,2011)。
付蓉蓉,劉楊,陳火英.2011.番茄黃化曲葉病的Ty-1和Ty-3抗性基因的PCR鑒定.分子植物育種(online),(9):1647-1652.
何自福,虞皓,毛明杰,羅方芳,林奕韓,王穗濤.2007.中國臺灣番茄曲葉病毒侵染引起廣東番茄黃化曲葉病.農業生物技術學報,15(1):119-123.
糾敏,周雪平,劉樹生.2006.煙粉虱傳播雙生病毒研究進展.昆蟲學報,49(3):513-520.
葉青靜,楊悅儉,王榮青,李志邈,阮美穎,周國治,姚祝平.2009.番茄抗黃化曲葉病育種研究進展.中國農業科學,42(4):1230-1242.
于力,朱龍英,萬延慧,楊少軍,朱為民,薛林寶.2008.多重PCR技術鑒定番茄Ty-1和Mi基因.分子植物育種,6(20):165-169.
余文貴,趙統敏,楊瑪麗,趙麗萍.2009.番茄黃化曲葉病及其抗病育種研究進展.江蘇農業學報,25(4):925-930.
趙統敏,余文貴,周益軍,季英華.2007.江蘇省番茄黃化曲葉病毒病(TYLCV)的發生與診斷初報.江蘇農業學報,23(6):654-655.
周雪平,崔曉峰,陶小容.2003.雙生病毒—— 一類值得重視的植物病毒.植物病理學報,33(6):487-492.
Anbinder I,Reuveni M,Azari R,Paran I,Nahon S,Shlomo H,Chen L,Lapidot M,Levin I.2009.Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum.Theor Appl Genet,119(3):519-530.
Hanson P M,Green S K,Kuo G.2006.Ty-2 a gene on chromosome 11 conditioning geminivirus resistance in tomato.Tomato Genetic Cooperative Report,56:17-18.
Ji Y F,Schuster D J,Scott J W.2007.Ty-3,a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato.Mol Breeding,20(3):271-284.
Ji Y F,Scott J W,Schuster D J,Maxwell D P.2009.Molecular mapping of Ty-4,a new Tomato yellow leaf curl virus resistance locus on chromosome 3 of tomato.Journal of the American Society for Horticultural Science,134(2):281-288.
Pico B,Diezm J,Nuez F.1996.Viral diseases causing the greatest economic losses to the tomato cropⅡ.The Tomato yellow leaf curl virus:a review.Sci Hortic(msterdam),67(3-4):151-196.
Vdavski F,Czosnek H,Gazit S.2008.Pyramiding of genes conferring resistance to Tomato yellow leaf curl virus from different wild tomato species.Plant Breeding,127:625-631.
Williamson V M,Ho J Y,Wu F F,Miller N,Kaloshian I.1994.A PCR-based marker tightly linked to the nematode resistance gene,Mi,in tomato.Theor Appl Genet,87:757-763.
Zamir D,Eksteinmi C,Heison I,Zakay Y.1994.Mapping and introgression of a Tomato yellow leaf curl virus tolerance gene,Ty-1.Theor Appl Genet,88(2):141-146.
