LIN28B在結(jié)腸癌中的表達及其臨床意義
2012-04-28 13:03:32龐明輝董丹丹梁耀澤胡李國新
首都醫(yī)科大學學報 2012年3期
龐明輝 王 康 伍 剛 張 偉 董丹丹 帥 平 梁耀澤胡 陽 李 平 李國新*
(1.南方醫(yī)科大學南方醫(yī)院普外科,廣州 510515;2.四川省人民醫(yī)院普外科,成都 610072;3.四川省人民醫(yī)院病理科,成都 610072;4.四川省人民醫(yī)院體檢中心,成都 610072)
LIN28參與形成誘導成體干細胞,它高表達于胚胎干細胞,隨細胞分化而逐漸低表達于分化細胞和成體組織。作為lin28的同源基因,lin28b在肝細胞癌中首先得到克隆,能夠在肝細胞癌中高表達并和細胞過度增生相關(guān)[1]。目前,lin28b被認為是“癌胚基因”,可促進細胞惡性轉(zhuǎn)化并特異高表達于部分進展期腫瘤,如肝細胞癌和結(jié)腸癌[2]。國內(nèi)尚無lin28b表達和結(jié)腸癌關(guān)系的報道。本研究利用EnVision免疫組織化學技術(shù)檢測美國Biomax結(jié)腸癌組織芯片(208例結(jié)腸癌,8例正常組織)和南方醫(yī)科大學南方醫(yī)院149例手術(shù)切除石蠟切片中l(wèi)in28b的表達并進行染色評分,行χ2檢驗研究其表達強度與結(jié)腸癌臨床病理特征的關(guān)系,判斷l(xiāng)in28b在結(jié)腸癌中的臨床價值。
1 材料與方法
1.1 研究對象
1)第一組研究對象:購買西安超英生物公司代理美國Biomax的結(jié)腸癌及正常組織組合微陣列芯片(編號CO2161),包含208例腫瘤和8例正常結(jié)腸組織的病理信息。該組織芯片樣點直徑1.0mm,厚度5 μm,采用中性甲醛固定。208例結(jié)腸癌中男118例,女90例,年齡24~82歲,平均年齡54.8歲,其高分化腺癌83例,中分化腺癌8例,低分化腺癌4例。Ⅰ期2例,Ⅱ期130例,Ⅲ期46例,Ⅳ期11例。發(fā)生淋巴結(jié)轉(zhuǎn)移55例,無淋巴結(jié)轉(zhuǎn)移153例,伴遠處轉(zhuǎn)移11例,無遠處轉(zhuǎn)移197例。詳見表1。
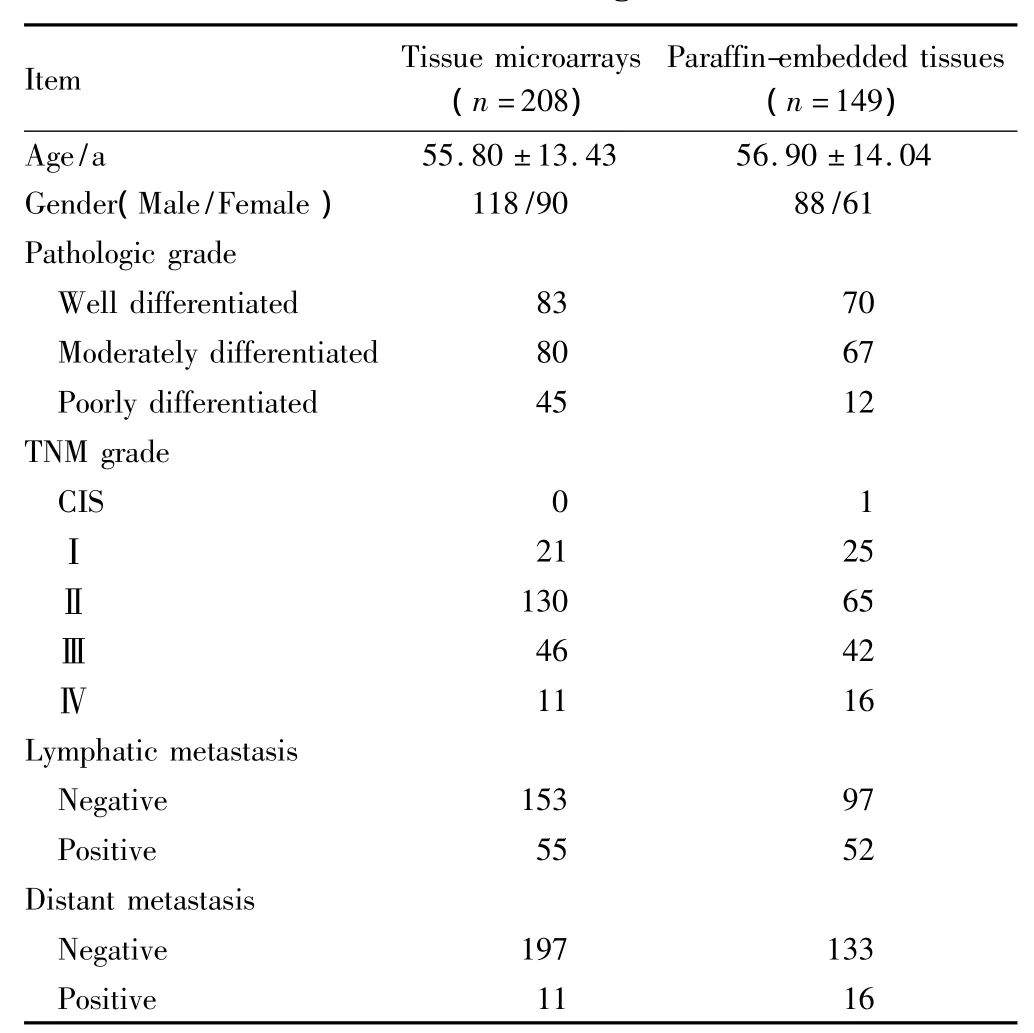
表1 臨床資料Tab.1 Clinical characteristics of patients/tumors at the time of diagnosis
2)第二組研究對象:在臨床研究倫理學會委員會批準、病人知情同意的情況下,選擇在南方醫(yī)科大學南方醫(yī)院普通外科2003年1月至2004年12月行手術(shù)切除的149例結(jié)腸癌組織標本,10%甲醛固定后石蠟包埋。所有標本均詳細記載病人基本信息,記錄腫瘤TNM分期、分化程度、淋巴結(jié)轉(zhuǎn)移和遠處轉(zhuǎn)移情況。其中,男性患者88例,女性61例,年齡19~85歲,平均年齡56.9歲;高分化腺癌70例,中分化腺癌67例,低分化腺癌12例,原位癌1例;TNMⅠ期25例,Ⅱ期65例,Ⅲ期42例,Ⅳ期16例;發(fā)生淋巴結(jié)轉(zhuǎn)移52例,無淋巴結(jié)轉(zhuǎn)移97例,遠處轉(zhuǎn)移16例,無遠處轉(zhuǎn)移133例。兩組研究對象均將高、中分化腺癌歸入分化較好組,將低分化腺癌歸入分化較差組。
3)入組與排除標準:
(1)入組標準:①首次因結(jié)腸癌就診,未經(jīng)治療病例;②結(jié)腸單一部位病變;③兩位以上病理醫(yī)師共同確診;④結(jié)腸癌病例有完整的隨訪資料;⑤術(shù)前均未行新輔助放化療。
(2)排除標準:①非首次就診病例;②結(jié)腸多部位病變病例;③家族性腺瘤性息肉及其他結(jié)腸疾病,例如潰瘍性結(jié)腸炎、結(jié)腸結(jié)核、克隆病等病例;④患有身體其他部位惡性腫瘤的病例;⑤有身體免疫缺陷疾病,例如人類免疫缺陷病毒(human immunodeficiency virus,HIV)攜帶者或獲得性免疫缺陷綜合征(acquired immunodeficiency syndrome,AIDS)患者的病例;⑥術(shù)前行新輔助放化療者。
1.2 免疫組織化學檢測
采用EnVision免疫組化技術(shù)(試劑盒購自丹麥Dako公司)檢測組織芯片和石蠟切片中l(wèi)in28b的表達。lin28b一抗購自英國Abcam公司,貨號ab71415,稀釋濃度1∶10,以PBS液代替一抗作陰性對照。由兩名病理科醫(yī)師在雙盲情況下對每個標本進行染色評分。結(jié)果按照如下標準進行判斷:陰性為無色,陽性為細胞質(zhì)染色呈棕黃色。根據(jù)陽性染色細胞百分比計分:高倍鏡下每張切片選擇10個有代表性的視野,每個視野計數(shù)100個腫瘤細胞共計數(shù)1000個細胞。0分為陽性細胞數(shù)小于或等于5%,1分為陽性細胞數(shù)10%~25%,2分為陽性細胞數(shù)25%~50%,3分為陽性細胞數(shù)大于50%,將0~2分計為低表達,3分為高表達[3]。
1.3 統(tǒng)計學方法
采用SPSS17.0統(tǒng)計軟件進行統(tǒng)計分析,計數(shù)資料的比較采用χ2檢驗。對單元格的期望頻數(shù)均大于1,但有小于1/5的單元格期望頻數(shù)小于5,采用連續(xù)性校正χ2檢驗。采用逐步回歸法進行多因素Logistic回歸分析。P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
1)美國Biomax結(jié)腸癌組織芯片免疫組化結(jié)果顯示,lin28b在正常結(jié)腸黏膜組織中均未染色,但在結(jié)腸癌組織中均有染色且隨分化程度的降低,lin28b表達強度增加(圖1)。
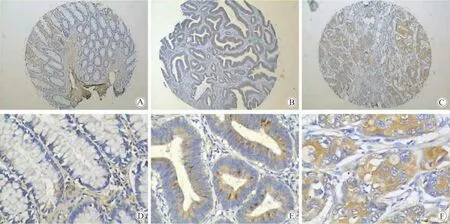
圖1 Lin28在美國biomax的組合芯片中的表達(免疫組化染色)Fig.1 Expression Lin28b in colon cancer tissue microarrays detected by immunohistochemistry
2)采用χ2檢驗分析lin28b表達強弱和結(jié)腸癌病理學特征之間的聯(lián)系,結(jié)果證實lin28b表達水平與腫瘤較差分化、較高TNM分期、淋巴結(jié)轉(zhuǎn)移及遠處轉(zhuǎn)移有關(guān)(P<0.001),見表2。
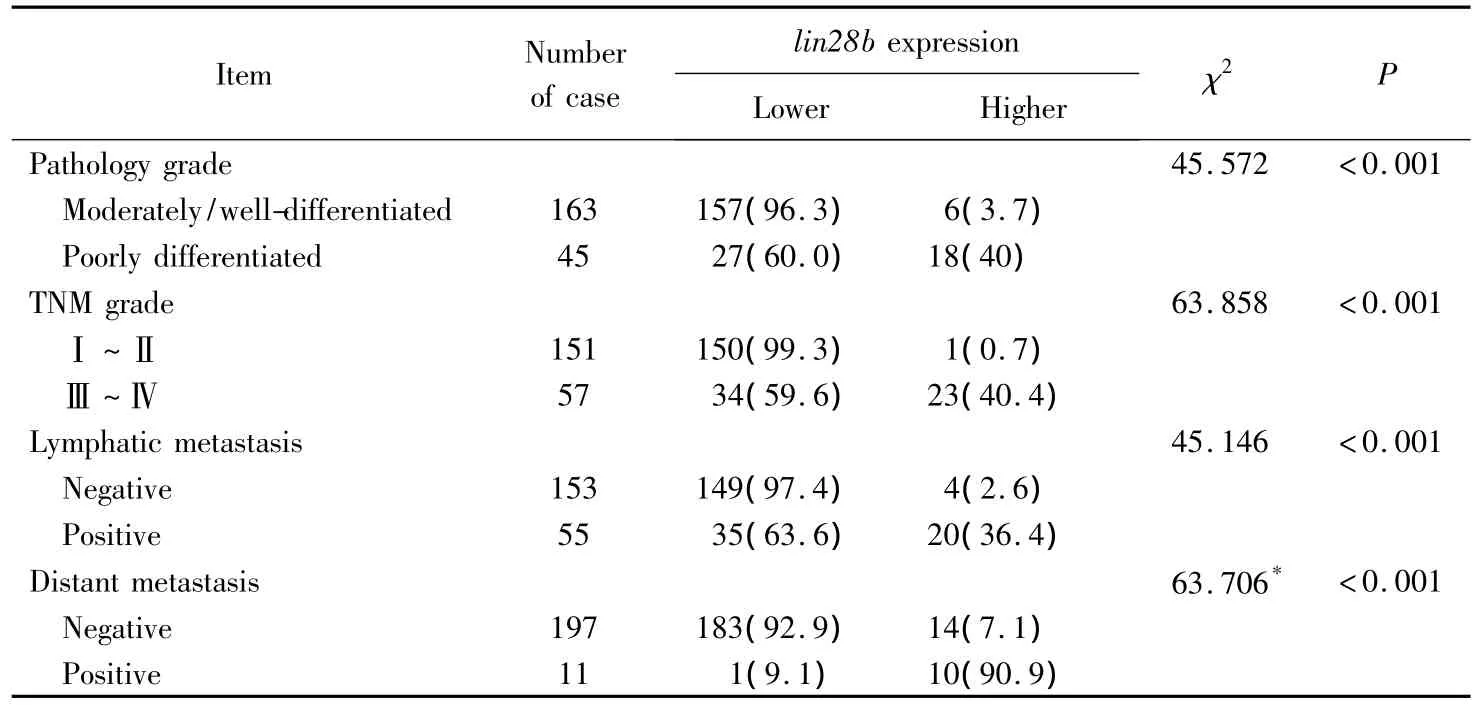
表2 組織芯片lin28b表達與結(jié)腸癌臨床病理之間的關(guān)系Tab.2 Expression of lin28b and its relationship with clinical pathology in colon cancer tissue microarrays n(%)
3)為驗證前述美國Biomax結(jié)腸癌組織芯片實驗的可重復性,本研究組進一步在中國南方醫(yī)院的結(jié)腸癌石蠟標本中重復上述實驗并得到一致的結(jié)果,顯示lin28b的表達在西方和中國人群中均與進展期結(jié)腸癌病理學特征(腫瘤較差分化、TNM分期Ⅲ~Ⅳ期、淋巴結(jié)轉(zhuǎn)移及遠處轉(zhuǎn)移)有關(guān),見表3。
4)由于組織芯片來源于美國Biomax公司,獲取的臨床信息有限,且無隨訪資料,所以我們僅對南方醫(yī)院收集到的149例臨床資料及隨訪信息完整的結(jié)腸癌組織切片的資料進行了Logistic多因素回歸分析,將lin28b表達作為因變量,年齡、性別、組織分化程度、TNM分期作為自變量納入回歸方程。逐步回歸分析結(jié)果顯示,僅TNM分期是lin28b高表達的獨立危險因素(OR=6.583,95%CI:2.97~14.59)。

表3 組織切片lin28b表達與結(jié)腸癌臨床病理之間的關(guān)系Tab.3 Expression of lin28b and its relationship with clinical pathology in paraffin-embedded surgically resected colon cancer tissues n(%)
3 討論
lin28基因編碼一種具有冷休克結(jié)構(gòu)域和反轉(zhuǎn)錄病毒型鋅指結(jié)構(gòu)的胞漿蛋白,在秀麗隱桿線蟲中最先發(fā)現(xiàn),參與維持干細胞“干性”。lin28b是lin28的同源基因,位于6q21,長度為752個堿基,GeneBank編號為NM_001004317。二者編碼的蛋白功能相近,在胚胎干細胞和發(fā)育組織中高表達,隨細胞分化而逐漸低表達,在腫瘤組織中則反常高表達。lin28b實質(zhì)為一種高度保守的RNA結(jié)合蛋白,大小為27 000,主要通過抑制非編碼的Let-7 microRNA家族來實現(xiàn)其促癌作用[4-7]。lin28b/Let-7通路與多種細胞生物學功能有關(guān),與多種腫瘤發(fā)生和發(fā)展相關(guān),其中包括結(jié)腸癌[8]。
國內(nèi)外多項研究認為lin28b多高表達于分化程度低的腫瘤[9-11]。本課題組研究結(jié)果顯示lin28b在正常結(jié)腸組織中未表達,在低分化結(jié)腸腺癌組織中表達較強,高分化組織中表達較弱,這說明隨腫瘤組織分化程度和lin28b表達存在負相關(guān)。Saiki Y等[12]研究發(fā)現(xiàn)lin28表達與結(jié)直腸癌淋巴結(jié)轉(zhuǎn)移及Dukes分期顯著相關(guān),本課題組的結(jié)果也證實Ⅲ、Ⅳ期結(jié)腸癌組織中l(wèi)in28b蛋白的表達顯著高于Ⅰ、Ⅱ期結(jié)腸癌。多因素Logistic回歸分析顯示TNM分期是lin28b高表達的獨立危險因素。因此,可認為結(jié)腸癌TNM分期越晚,lin28b的表達越強。
轉(zhuǎn)移和復發(fā)是結(jié)腸癌患者死亡的主要原因,轉(zhuǎn)移發(fā)生包括細胞與基底膜黏附、細胞外基質(zhì)的降解及細胞的遷出進入血管[13]。Wang Y C 等[14]研究認為lin28b高表達可增加肝癌細胞侵襲能力,蔡民等[15]證實lin28b/Let-7通路對膽囊癌細胞在降解細胞外基質(zhì)及隨后的遷移方面有著重要作用。Klemke M等[16]研究指出lin28通過抑制Let-7合成并上調(diào)Let-7下游癌基因hmga2的表達而最終促進上皮細胞向間充質(zhì)細胞轉(zhuǎn)化,對腫瘤轉(zhuǎn)移發(fā)生具有重要意義[17]。本課題組研究結(jié)果顯示lin28b高表達與淋巴結(jié)轉(zhuǎn)移及遠處轉(zhuǎn)移有關(guān),提示lin28b高表達可能促進結(jié)腸癌細胞侵襲及轉(zhuǎn)移,下一步將采用Transwell實驗從分子水平研究lin28b對結(jié)腸癌細胞遷移及侵襲能力的影響。
綜上,本研究采用免疫組織化學法檢測干細胞相關(guān)因子lin28b在結(jié)腸癌中的表達,結(jié)果顯示lin28b的高表達與結(jié)腸癌低分化、TNM分期Ⅲ-Ⅳ期、淋巴結(jié)轉(zhuǎn)移及遠處轉(zhuǎn)移顯著相關(guān),其表達水平可作為判斷結(jié)腸癌患者病程進展及預后的重要指標,本研究結(jié)果有望為結(jié)腸癌靶向治療提供新的治療靶點。進一步我們將利用完善的隨訪資料進行l(wèi)in28b表達水平與結(jié)腸癌病人術(shù)后生存及復發(fā)方面的系列研究。
[1]Guo Y,Chen Y,Ito H,et al.Identification and characterization of lin-28 homolog B(LIN28B)in human hepatocellular carcinoma[J].Gene,2006,384(1):51-61.
[2]Viswanathan S R,Daley G Q.Lin28:A microRNA regulator with a macro role[J].Cell,2010,140(4):445-449.
[3]King C,Cuatrecasas M,Castells A,et al.LIN28B pro-motes colon cancer progression and metastasis[J].Cancer Res,2011,71(12):4260-4268.
[4]King C,Wang L,Winograd R,et al.LIN28B fosters colon cancer migration,invasion and transformation through let-7-dependent and-independent mechanisms[J].Oncogene,2011,30(40):4185-4193.
[5]Viswanathan S R,Daley G Q,Gregory R I.Selective blockade of microRNA processing by Lin28[J].Science,2008,320(5872):97-100.
[6]Rybak A,fuchs H ,Smirnova L,et al.A feedback loop comprising Lin-28 and let-7 conrols pre-let-7 maturation during neural stem-cell commitment[J].Nat Cell Biol,2008,10(8):987-993.
[7]Heo I,Joo C,Cho J,et al.Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA [J].Mol Cell,2008,32(2):276-284.
[8]Viswanathan SR,Powers JT,Einhorn W,et al.Lin28 promotes transformation and is associated with advanced human malignancies[J].Nat Genet,2009,41(7):843-848.
[9]Helland A,Anglesio MS,George J,et al.Deregulation of MYCN,LIN28B and LET7 in a molecular subtype of aggressive high-grade serous ovarian cancers[J].PLoS One,2011,6(4):e18064.
[10]Peng S,Maihle N J,Huang Y.Pluripotency factors Lin28 and Oct4 identify a sub-population of stem cell-like cells in ovarian cance[J].Oncogene,2010,29(14):2153-2159.
[11]羅玄,吳云林,馮立新.Lin28對胃腺癌細胞株增殖周期的影響及機制[J].內(nèi)科理論與實踐,2010,5(1):62-67.
[12]Saiki Y,Ishimaru S,Mimori K,et al.Comprehensive analysis of the clinical significance of inducing pluripotent stemness-related gene expression in colorectal cancer cells[J].Ann Surg Oncol,2009,16(9):2638-2644.
[13]關(guān)亞偉,李世擁,安萍,等.結(jié)直腸癌患者血清中組織因子含量與臨床意義的研究[J].華北國防醫(yī)藥,2009,21(2):1-3.
[14]Wang Y C,Chen Y L,Yuan R H et al.Lin-28B expression promotes transformation and invasion in human hepatocellular carcinoma[J].Carcinogenesis,2010,31(9):1516-1522.
[15]蔡民,李立波.靶向c-myc基因小RNA干擾對膽囊癌細胞侵襲運動的影響[J].第二軍醫(yī)大學學報,2010,31(9):1028-1031.
[16]Klemke M,Meyer A,Hashemi Nezhad M,et al.Loss of let-7 binding sites resulting from truncations of the 3'untranslated region of HMGA2 mRNA in uterine leiomyomas[J].Cancer Genet Cytogenet,2010,196(2):119-123.
[17]Dangi-Garimella S,Yun J,Eves E M ,et al.Raf kinase inhibitory protein suppresses a metastasis signalling cascade involving LIN28 and let-7[J].EMBO J,2009,28(4):347-358.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
西南國防醫(yī)藥(2016年7期)2016-12-01 06:01:15
中國衛(wèi)生標準管理(2015年1期)2016-01-14 03:41:26
河南醫(yī)學研究(2014年3期)2014-02-27 14:51:48
沈陽醫(yī)學院學報(2014年1期)2014-02-16 06:19:24
