多漿旱生植物霸王質(zhì)膜Na╋/H╋逆向轉(zhuǎn)運(yùn)蛋白基因RNAi載體構(gòu)建
2012-03-13 03:39:56王鎖民
草業(yè)科學(xué) 2012年4期
馬 清,王鎖民
(草地農(nóng)業(yè)生態(tài)系統(tǒng)國(guó)家重點(diǎn)實(shí)驗(yàn)室 蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院,甘肅 蘭州 730020)
霸王(Zygophyllumxanthoxylum)為蒺藜科(Zygophyllaceae)多漿旱生灌木,其抗逆性強(qiáng),是亞洲中部荒漠區(qū)的特有植物種,也是我國(guó)西部荒漠區(qū)植被組成的優(yōu)勢(shì)建群種,具有很高的生態(tài)和飼用價(jià)值[1-4]。Wang等[5]發(fā)現(xiàn)霸王適應(yīng)干旱環(huán)境的最有效策略是吸收并積累大量的Na+。進(jìn)一步分析發(fā)現(xiàn),干旱脅迫下,霸王存在著一種非常強(qiáng)大的長(zhǎng)距離運(yùn)輸能力,能夠?qū)⒏滴盏拇罅縉a+及時(shí)有效地轉(zhuǎn)運(yùn)至葉中,用以維持細(xì)胞的滲透勢(shì)[6-7],而質(zhì)膜Na+/H+逆向轉(zhuǎn)運(yùn)蛋白SOS1可能在植物體內(nèi)Na+的長(zhǎng)距離運(yùn)輸及空間分配中發(fā)揮重要作用[8-9]。為此,蘭州大學(xué)草類逆境生理與基因工程實(shí)驗(yàn)室從霸王中克隆了質(zhì)膜Na+/H+逆向轉(zhuǎn)運(yùn)蛋白基因ZxSOS1(Genebank登錄號(hào):GU177864),但ZxSOS1在霸王體內(nèi)Na+轉(zhuǎn)運(yùn)中的功能及其在霸王適應(yīng)干旱生境中的作用機(jī)制尚不清楚。
在基因功能研究中,RNA干擾技術(shù)(RNA interference,RNAi)已成為一種簡(jiǎn)單、高效、特異和成本相對(duì)低廉的研究手段,它可以在mRNA水平上使特定基因的表達(dá)活性喪失或下降,從而獲得功能性突變或喪失,進(jìn)而可根據(jù)表型變化鑒定目標(biāo)基因的功能[10-11]。因此,本研究構(gòu)建以霸王ZxSOS1基因?yàn)榘袠?biāo)的、具有反向重復(fù)發(fā)卡結(jié)構(gòu)的RNAi表達(dá)載體,旨在為利用RNAi技術(shù)深入研究ZxSOS1在霸王體內(nèi)Na+長(zhǎng)距離運(yùn)輸及空間分配中的功能及其作用機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1材料
1.1.1試驗(yàn)材料 霸王種子于2006年7月采自內(nèi)蒙古阿拉善左旗巴彥浩特。pHANNIBAL質(zhì)粒、PART27質(zhì)粒和大腸桿菌E.coliDH5α菌株均為蘭州大學(xué)草類逆境生理與基因工程實(shí)驗(yàn)室保存,引物由上海生工生物工程公司合成。
1.1.2試劑 UNIQ-10柱式Trizol總RNA抽提試劑盒、MMLV第一鏈cDNA合成試劑盒、PCR擴(kuò)增試劑盒和UNIQ-10柱式DNA膠回收試劑盒均購(gòu)自上海生工生物工程公司,克隆載體pGM18-T Easy vector、限制性內(nèi)切酶和T4 DNA連接酶均購(gòu)自TaKaRa公司,DNA marker購(gòu)自北京天根生化科技有限公司,其他生化試劑均為進(jìn)口或國(guó)產(chǎn)分析純。
1.2試驗(yàn)方法
1.2.1總RNA的提取 取50 mmol·L-1NaCl處理24 h的3周齡霸王幼苗根系,加入液氮研磨至粉末狀,按照UNIQ-10柱式Trizol總RNA抽提試劑盒的操作說(shuō)明書提取根系總RNA。用1.0%甲醛變性凝膠電泳鑒定其完整性和質(zhì)量。
1.2.2ZxSOS1基因RNAi靶位片段的擴(kuò)增 cDNA第一鏈的合成按照MMLV第一鏈cDNA合成試劑盒的操作說(shuō)明進(jìn)行。
利用GenBank中的RNAi設(shè)計(jì)庫(kù)對(duì)ZxSOS1基因的靶位區(qū)段進(jìn)行篩選,選取629 bp(第2 750~3 379位堿基)區(qū)段作為RNAi最佳靶位區(qū)段設(shè)計(jì)一對(duì)特異性引物,為了便于定向連接到中間載體pHANNIBAL,在上游引物P1上引入酶切位點(diǎn)Xba Ⅰ和Xho Ⅰ序列,下游引物P2上引入酶切位點(diǎn)Hind Ⅲ和Kpn Ⅰ序列。具體引物序列如下(劃線部分為酶切位點(diǎn)序列):
P1:5′-TCTAGACTCGAGTCAACAATGACTGTATACTT-3′;
P2:5′-AAGCTTGGTACCAATTTCCTCAAATCACGCTT-3′。
PCR擴(kuò)增反應(yīng)體系:10×PCR Buffer 5 μL,25 mmol·L-1MgCl23 μL,2 mmol·L-1dNTP 5 μL,10 μmol·L-1P1 1 μL,10 μmol·L-1P2 1 μL,Taq DNA polymerase (5 U· μL-1)0.5 μL,cDNA 2 μL,加水至50 μL。反應(yīng)條件為94 ℃預(yù)變性2 min;94 ℃變性30 s、53 ℃退火45 s、72 ℃延伸50 s,30個(gè)循環(huán);最后72 ℃延伸10 min。
PCR擴(kuò)增產(chǎn)物用1.0%瓊脂糖凝膠檢測(cè),目的片斷的回收和純化按照UNIQ-10柱式DNA膠回收試劑盒操作說(shuō)明進(jìn)行。回收的PCR產(chǎn)物連接到pGM18-T Easy vector載體,轉(zhuǎn)化感受態(tài)大腸桿菌E.coliDH5α,用含有50 g·mL-1氨芐青霉素的LB固體培養(yǎng)基進(jìn)行藍(lán)白斑篩選,陽(yáng)性克隆經(jīng)質(zhì)粒PCR鑒定確認(rèn)后,送至華大基因科技股份有限公司進(jìn)行基因測(cè)序,使用DNAMAN生物軟件對(duì)序列進(jìn)行分析。
1.2.3以ZxSOS1為靶標(biāo)的RNAi載體構(gòu)建 用Xho /ⅠKpn Ⅰ雙酶切pGM18-T Easy vector載體,回收小片段(S1),與經(jīng)同樣雙酶切并回收的載體pHANNIBAL連接,轉(zhuǎn)化感受態(tài)E.coliDH5α,得到重組質(zhì)粒pHS1;然后用Xba Ⅰ/Hind Ⅲ雙酶切pGM18-T Easy vector載體并回收反向小片段(S2),與經(jīng)同樣雙酶切并回收的pHS1載體相連,得到以pHANNIBAL載體的內(nèi)含子(intron)部位作為間隔區(qū)的、具有發(fā)卡反向重復(fù)序列的中間載體pHS1S2(圖1);最后用Sac Ⅰ/Spe Ⅰ雙酶切pHS1S2,回收大片段,與經(jīng)同樣酶切回收的PART27載體相連,得到具有ZxSOS1反向重復(fù)序列的植物RNAi表達(dá)載體PARS(圖2)。
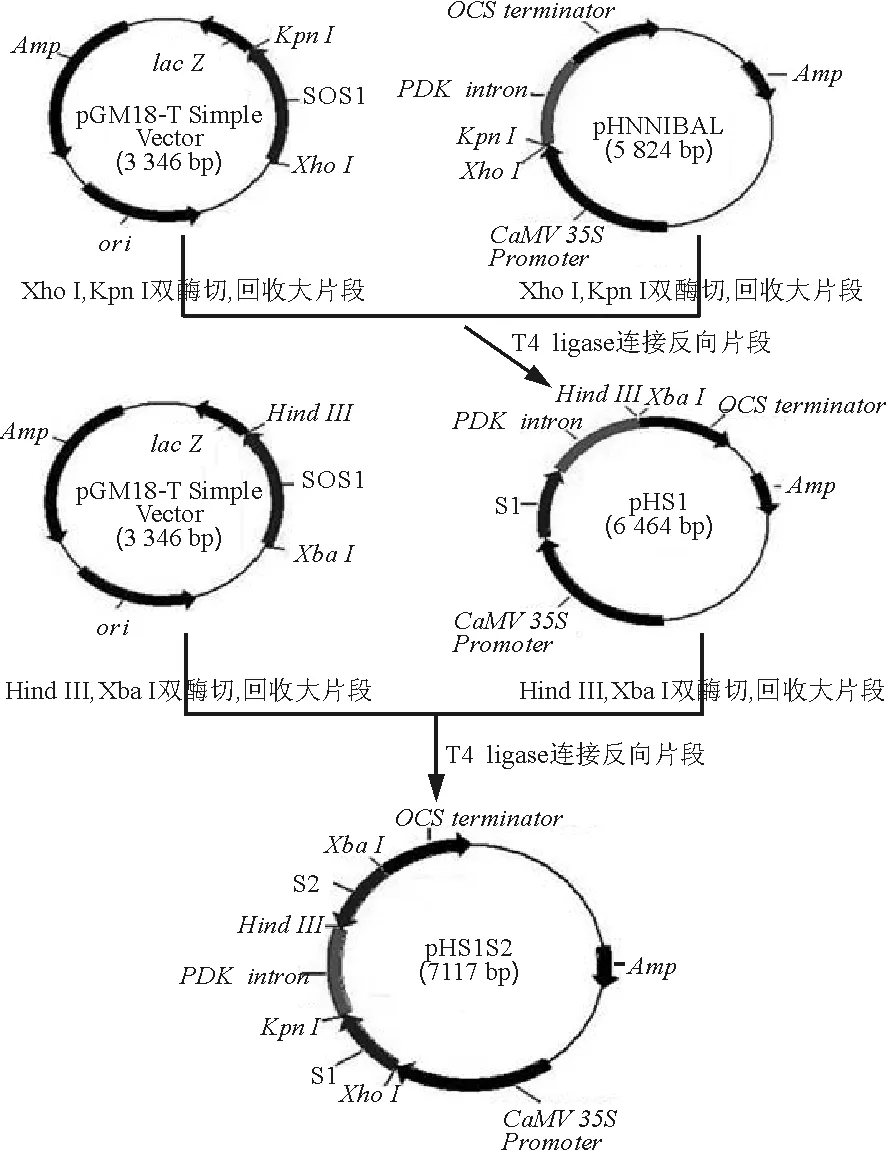
圖1 中間載體pHS1S2的構(gòu)建
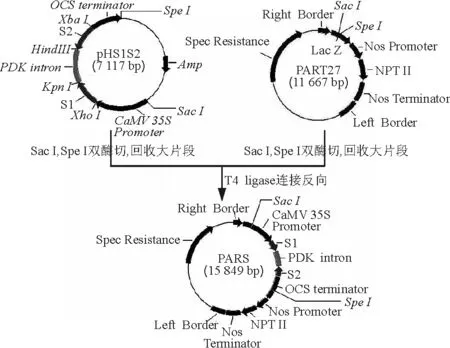
圖2 ZxSOS1 RNAi載體PARS的構(gòu)建
2 結(jié)果與分析
2.1總RNA的提取及檢測(cè) 以霸王根系為材料提取總RNA,甲醛變性凝膠電泳檢測(cè)結(jié)果顯示28 S rRNA、18 S rRNA條帶清晰(圖3),前者亮度約是后者的2倍,說(shuō)明所提取的RNA完整性較好;經(jīng)NANODROP1000核酸蛋白檢測(cè)儀測(cè)得OD260 nm/OD280 nm平均值為2.01,表明RNA純度較高,可以用于RT-PCR擴(kuò)增實(shí)驗(yàn)。
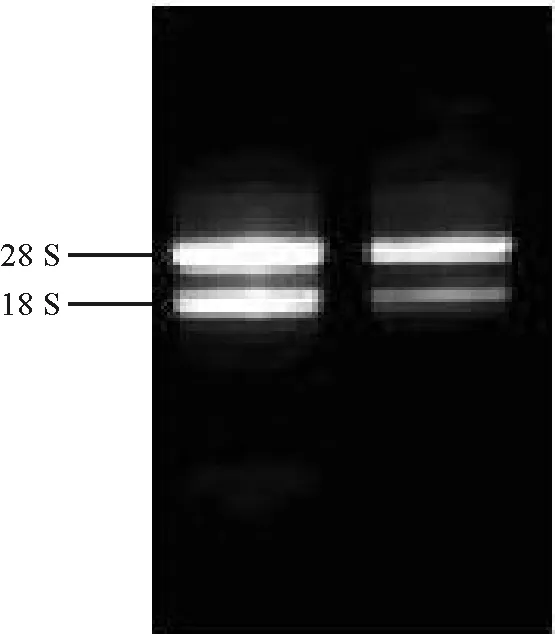
圖3 霸王根系總RNA的甲醛變性凝膠電泳圖
2.2ZxSOS1基因靶位片段的確定及擴(kuò)增 利用GenBank中的RNAi 設(shè)計(jì)庫(kù)對(duì)ZxSOS1基因的靶位區(qū)段進(jìn)行篩選,結(jié)果表明,ZxSOS1基因從起始密碼開始第2 750~3 379位堿基共629 bp的片段為保守區(qū)段。以總RNA反轉(zhuǎn)錄所得到的第一鏈cDNA為模板,用特異引物P1和P2進(jìn)行PCR擴(kuò)增,得到了預(yù)期大小的特異條帶(圖4)。將此條帶回收純化后連接到pGM18-T Easy vector載體上,轉(zhuǎn)化E.coliDH5α。從轉(zhuǎn)化的平板上隨機(jī)挑取3個(gè)菌斑并提取質(zhì)粒,進(jìn)行PCR擴(kuò)增,得到了629 bp的預(yù)期條帶(結(jié)果未顯示),表明這些克隆為陽(yáng)性克隆。對(duì)其進(jìn)行測(cè)序后分析表明,該序列與ZxSOS1基因相應(yīng)區(qū)段同源性100%(結(jié)果未顯示),說(shuō)明已成功擴(kuò)增到目的片段。
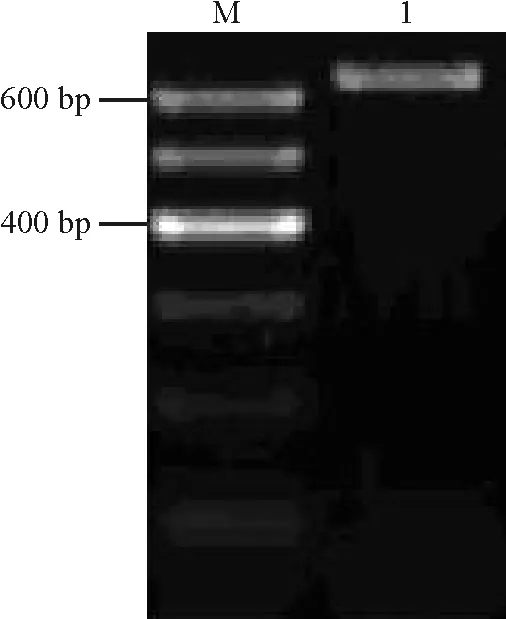
圖4 RT-PCR產(chǎn)物凝膠電泳圖
2.3以ZxSOS1為靶標(biāo)的RNAi載體構(gòu)建 按照Z(yǔ)xSOS1 RNAi植物表達(dá)載體的構(gòu)建流程(圖1、圖2),通過(guò)酶切和連接的方法,構(gòu)建了具有ZxSOS1反向重復(fù)序列的植物RNAi表達(dá)載體PARS。通過(guò)PCR檢測(cè)鑒定出PARS的陽(yáng)性重組質(zhì)粒后(圖5),根據(jù)PARS載體上的限制性位點(diǎn)對(duì)其進(jìn)行如下酶切分析:經(jīng)Xho Ⅰ/Kpn Ⅰ雙酶切后,在800和629 bp處出現(xiàn)特異條帶(圖6,泳道1),分別與PDK intron和S1片段大小相符;經(jīng)Xba Ⅰ/Hind Ⅲ雙酶切后,在629 bp處出現(xiàn)特異條帶(圖6,泳道2),與S2片段大小相符;經(jīng)Sac Ⅰ/Spe Ⅰ雙酶切后,在約4 500 bp處出現(xiàn)特異條帶(圖6,泳道3),與CaMV 35S啟動(dòng)子、S1、PDK intron、S2及OCS終止子片段之和大小相符。以上結(jié)果表明,以ZxSOS1為靶標(biāo)的RNAi載體已經(jīng)構(gòu)建成功。
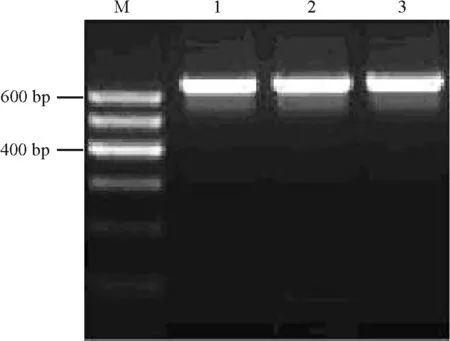
圖5 PARS重組質(zhì)粒陽(yáng)性克隆的PCR鑒定
3 討論與結(jié)論
生長(zhǎng)在荒漠地區(qū)的多漿旱生植物霸王具有極強(qiáng)的抗逆性,可能蘊(yùn)藏著豐富的抗逆基因資源[7]。基因沉默是研究基因功能的常用手段,包括反義核酸、基因敲除和RNAi等技術(shù)[12]。相比其他基因沉默方法,RNAi具有高效性,只需要少量小干擾RNA(small-interference RNA,siRNA)或雙鏈RNA(double-stranded RNA,dsRNA)即可使特定基因沉默[11,13];且沉默效應(yīng)能通過(guò)植物胞間連絲及韌皮部進(jìn)行系統(tǒng)擴(kuò)散[14-19];同時(shí)植物中的RNAi沉默效應(yīng)可以從起始siRNA同源區(qū)向臨近的5′和3′端擴(kuò)散,進(jìn)而可增強(qiáng)基因沉默的效果[20]。因此,RNAi已逐漸成為一種研究植物基因功能的強(qiáng)有力工具。
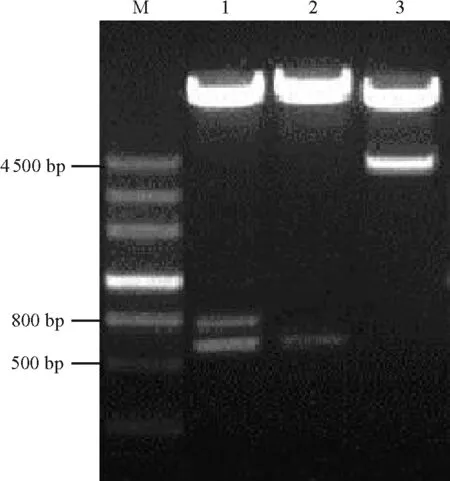
圖6 PARS的酶切分析
RNAi的關(guān)鍵是其載體的構(gòu)建,載體結(jié)構(gòu)、靶基因序列長(zhǎng)度及其在靶基因中的位置將直接影響基因沉默的效果[21]。RNAi載體以含有2個(gè)反向重復(fù)序列中間夾1個(gè)內(nèi)含子所形成ihpRNA發(fā)卡結(jié)構(gòu)的沉默效果最好,平均比例較高,超過(guò)90%[22],其發(fā)夾結(jié)構(gòu)臂序列的長(zhǎng)度具有一定的可塑性,一般在100~1 200 bp[23]。Elbashir等[24]和Reynolds等[25]建議靶基因序列應(yīng)在基因轉(zhuǎn)錄起始位點(diǎn)下游100個(gè)核苷酸以后,且該段DNA的GC含量在40%~52%。本研究構(gòu)建的RNAi載體的2個(gè)反向重復(fù)序列臂之間具有內(nèi)含子,且兩臂大小也在目前公認(rèn)的序列長(zhǎng)度范圍之內(nèi),選取的ZxSOS1基因靶序列GC含量為44%。因此,該載體完全滿足高效RNAi載體的條件,理論上將會(huì)對(duì)ZxSOS1基因產(chǎn)生較好的沉默效果。
[1]趙一之,朱宗元.亞洲中部荒漠區(qū)的植物特有屬[J].云南植物研究,2003,25(2):113-121.
[2]楊鑫光,傅華,牛得草.干旱脅迫下幼苗期霸王的生理響應(yīng)[J].草業(yè)學(xué)報(bào),2007,16(5):107-112.
[3]周向睿,周志宇,吳彩霞.霸王繁殖特性的研究[J].草業(yè)科學(xué),2006,23(6):38-41.
[4]吳彩霞,周志宇,莊光輝,等.強(qiáng)旱生植物霸王和紅砂地上部營(yíng)養(yǎng)物質(zhì)含量及其季節(jié)動(dòng)態(tài)[J].草業(yè)科學(xué),2004,21(3):30-34.
[5]Wang S M,Wan C G,Wang Y R,etal.The characteristics of Na+,K+and free proline distribution in several drought resistant plants of the Alxa Desert,China[J].Journal of Arid Environments,2004,56:525-539.
[6]李三相.Na+與多漿旱生植物霸王抗旱性研究[D].蘭州:蘭州大學(xué),2006.
[7]Wu G Q,Xi J J,Wang Q,etal.The ZxNHX gene encoding tonoplast Na+/H+antiporter from the xerophyteZygophyllumxanthoxylumplays important roles in response to salt and drought[J].Journal of Plant Physiology,2011,168:758-767.
[8]Shi H,Quintero F J,Pardo J M,etal.The putative plasma membrane Na+/H+antiporter SOS1 controls long distance Na+transport in plants[J].Plant Cel1,2002,14:465-477.
[9]Olías R,Eljakaoui Z,Li J,etal.The plasma membrane Na+/H+antiporter SOS1 is essential for salt tolerance in tomato and affects the partitioning of Na+between plant organs[J].Plant,Cell and Environment,2009,32:904-916.
[10]Fire A,Xu S,Montgomery M K,etal.Potent and specific genetic interference by double-stranded RNA inCaenorhabditiselegans[J].Nature,1998,391:806-811.
[11]Meister G,Tuschl T.Mechanisms of gene silencing by double-stranded RNA[J].Nature,2004,431:343-349.
[12]殷勤偉.siRNA介導(dǎo)的基因沉默[J].生物物理學(xué)報(bào),2004,20(2):85-95.
[13]Baulcombe D.RNA silencing in plants[J].Nature,2004,431:356-363.
[14]Mallory A C,Mlotshwa S,Bowman L H,etal.The capacity of transgenic tobacco to send a systemic RNA silencing signal depends on the nature of the inducing transgene locus[J].The Plant Journal,2003,35:82-92.
[15]Palauqui J C,Balzergue S.Activation of systemic acquired silencing by localised introduction of DNA[J].Current Biology,1999,9:59-66.
[16]Palauqui J C,Elmayan T,Pollien J M,etal.Systemic acquired silencing: transgene-specific post- transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions[J].The EMBO Journal,1997,16:4738-4745.
[17]Sonoda S,Nishiguchi M.Graft transmission of post-transcriptional gene silencing: target specificity for RNA degradation is transmissible between silenced and non-silenced plants, but not between silenced plants[J].The Plant Journal,2000,21:1-8.
[18]Tournier B,Tabler M,Kalantidis K.Phloem flow strongly influences the systemic spread of silencing in GFPNicotianabenthamianaplants[J].The Plant Journal,2006,47:383-394.
[19]Voinnet O,Vain P,Angell S.Systemic spread of sequence-specific transgene RNA degradation in plants is initiated by localized introduction of ectopic promoterless DNA[J].Cell,1998,95:177-187.
[20]白描,楊國(guó)順,陳石,等.植物RNAi的特點(diǎn)及其應(yīng)用研究進(jìn)展[J].生物技術(shù)通報(bào),2009(8):6-10.
[21]許德暉,黃辰,劉利英,等.高效siRNA設(shè)計(jì)的研究進(jìn)展[J].遺傳,2006,28(11):1457-1461.
[22]Wesley S V,Helliwell C A,Smith N A,etal.Construct design for efficient, effective and high-throughput gene silencing in plants[J].The Plant Journal,2001,27:581-590.
[23]Hirai S,Oka S,Adachi E,etal.The effect s of spacer sequences on silencing efficiency of plant RNAi vectors[J].Plant Cell Reports,2007,26:651-659.
[24]Elbashir S M,Lendeckel W,Tuschl T.RNA interference is mediated by 21- and 22- nucleotide RNAs[J].Genes and Development,2001,15:188-200.
[25]Reynolds A,Leake D,Boese Q,etal.Rational siRNA design for RNA interference[J].Nature Biotechnology,2004,22:326-330.
