大豆食心蟲28SrDNA基因的克隆及表達分析
2012-03-12 06:21:42李冬梅孟凡立王志坤臧振源李文濱
東北農業大學學報 2012年7期
李冬梅,孟凡立,王志坤,臧振源,孫 晶,李文濱
(東北農業大學農學院,大豆生物學教育部重點實驗室,哈爾濱 150030)
大豆食心蟲(Leguminivora glycinivorella Matsumurav)屬鱗翅目(Lepidoptera)小卷蛾科(Olethreutidae),是東北大豆產區最嚴重的一種害蟲,以幼蟲鉆蛀豆莢取食豆粒,將豆粒咬成溝道或殘破狀,嚴重影響大豆產量和品質[1]。豆農每年至少使用2次化學殺蟲劑對大豆食心蟲進行防治。現在使用的化學殺蟲劑多為廣譜性殺蟲劑,可殺死目標昆蟲,也會殺死有益昆蟲。同時化學殺蟲劑在自然界可長期存在,在土壤和河流中逐漸積累,破壞土壤和河流的生態環境;化學殺蟲劑還可以通過食物鏈進入人體,誘發致畸、致癌,嚴重損害健康。利用傳統育種方法培育抗大豆食心蟲的大豆品種雖有效,但周期長、效率低。因而,急需探索防治大豆食心蟲的新方法,以精確、高效地控制大豆食心蟲。
Fire等在研究反義RNA在線蟲Caenorhabditis elagans中的基因阻斷作用時,首先發現利用純化的外源dsRNA能夠高效特異性阻斷內源基因表達,并將這一現象稱為 RNAi(RNA interference)[2]。由于RNAi技術沉默基因的高效性和特異性,人們提出通過轉基因手段,讓植物體內表達昆蟲基因的dsRNA,dsRNA經昆蟲取食后進入蟲體內,對昆蟲靶基因進行RNA干擾,從而特異性抑制靶基因表達,提高植物抗蟲性[3-6]。中國科學院上海生命科學研究院植物生理生態研究所陳曉亞院士實驗室的研究成果證明,昆蟲取食表達dsRNA的轉基因植物后,體內相應的靶基因受到抑制,提高植物的抗蟲性[7]。Mao等先通過轉基因手段,讓擬南芥和煙草表達棉鈴蟲參與棉毒素解毒的P450基因dsRNA。然后,利用轉基因植物喂食棉鈴蟲,dsRNA從食道進入細胞,抑制棉鈴蟲體內P450基因的表達,致使棉鈴蟲對棉子酚的抗性降低。最后,對棉鈴蟲造成致命的影響[8]。孟山都公司報道表明,玉米根蟲V-ATPase基因被轉基因玉米表達的siRNA抑制,轉基因玉米根部遭受的破壞要比對照植株顯著降低[9]。以上研究結果表明,dsRNA可以通過取食從植物傳遞到昆蟲體內,通過植物介導的RNAi技術可以干擾害蟲的重要基因,達到防治害蟲的目的,為利用RNAi技術培育新一代抗蟲轉基因作物提供重要的科學依據。
核糖體是普遍存在于各種細胞內的細胞器,由大小兩個亞基組成,是蛋白質合成的唯一場所。核糖體含有50%~60%的核糖體RNA(rRNA),其余為蛋白質。近年,國內外的研究表明,核糖體RNA(rRNA)不僅是構成核糖體的主要成分,還具有自我復制和自我剪切功能[10],在蛋白質合成中起重要作用[11]。28SrRNA是核糖體RNA主要成員之一,是生物的關鍵生長基因。因此,本試驗根據GenBank上已經發表的昆蟲28SrDNA基因序列,利用28SrDNA基因序列的保守區域,克隆大豆食心蟲28SrDNA序列全長,利用熒光定量PCR對大豆食心蟲不同生育時期和幼蟲的不同組織的28SrDNA表達量進行分析后,構建表達大豆食心蟲28SrDNA dsRNA的重組載體L4440-28SrDNA,為進一步研究RNA干擾技術防治大豆食心蟲奠定基礎。
[1]Laufer,B.&P.,Nation.(1999).A Vocabulary Size Test of Controlled Productive Ability,Language Testing,16(1).33-51.
1 材料與方法
1.1 材料
2010年7月末8月初大豆田間出現大豆食心蟲成蟲開始,收集卵、幼蟲、蛹、成蟲四個時期的大豆食心蟲樣品,將收集的蟲樣分別放入無Rnase的EP管中,立即放入液氮中冷凍,然后存于-80℃低溫冰箱,每個處理重復3次。選取大豆食心蟲3~4齡幼蟲進行解剖,剖取神經節、表皮、唾腺、中腸、卵巢、睪丸和脂肪體組織后,分別放入無Rnase的EP管中,立即放入-80℃低溫冰箱中保存。
DNA提取、PCR、電泳所用試劑均購自上海生工公司,測序由華大基因有限公司完成、OMEGA膠回收試劑盒、Trizol reagen試劑盒、MEGA4.0軟件、DNAman 5.2.2軟件、載體L4440和菌種HT115均由美國線蟲遺傳學中心提供。
嵌入式計算機在接到指定任務后,必須在規定時間內給出實時應答,這是區別嵌入式計算機與通用計算機的一個重要特征,實時性涉及到硬件的性能、軟件的中斷管理和調度算法等。
Design of Intelligent Balancing Vehicle Control System based on Arduino
2.3 術后硬膜下并發癥影響因素分析 單因素分析結果(表3)表明:男性、高齡、動脈瘤部位、Fisher分級、腦萎縮程度及術后腰大池引流是破裂動脈瘤夾閉術后硬膜下并發癥發生的潛在危險因素(P<0.05)。
1.2 方法
1.2.1 大豆食心蟲基因組DNA的提取
種植雙胞山藥最好選擇土質疏松、地勢高燥、土層深厚、排灌方便、通氣透水、保水保肥性能好的砂壤土。鹽堿地、低洼積水地、賣窯泥地、地下異物多的地不能種山藥。
縣級設專職人影管理人員1名,作業人員4~5名(高炮作業人員除外)。防雹作業點均交由當地政府管理,施行鄉鎮武裝部長主管、村干部管理、炮長負責的制度,每個作業點配備4名作業人員,一人為炮長,具體負責作業點管理、高炮維護、彈藥交接、作業實施及空域請示,氣象部門僅負責安全檢查、作業指導和空域請示等。
根據GenBank上已經發表鱗翅目昆蟲的銀月豹鳳蝶(Papilio troilus,登錄號AF423920);家蠶(Bombyx mori,登錄號M31320);雄鹿天蠶蛾(Hemileuca sp,登錄號AF423922);洋槐木蠹蛾(Prionoxystus robiniae,登錄號AY521785)的28SrDNA序列,根據鱗翅目昆蟲28SrDNA序列的保守區域設計兩對簡并引物用于擴增大豆食心蟲28SrDNA序列全長,引物具體如下:
利用DNAMAN5.2.2和MEGA4.0軟件進行序列分析。
P2:ATGGTGAACTATGCCTGGT;
P3:AACTTCGGAGGGAACCG;
P4:CCCTCCCTGRATTTCAAGGTCCG。
按昆蟲DNA提取方法,利用試劑盒提取大豆食心蟲幼蟲DNA。
P1和P2預期的PCR擴增條帶大小約為1100 bp,P3和P4約為1200 bp。
1.2.3 PCR擴增與檢測
PCR反應體系為PCR Buffer 2.5 μL,TaqDNA聚合酶 0.3 μL,dNTPs 0.5 μL,10 mmol·L-1引物 1.0 μL,模板DNA 2.5 U 1.0 μL,ddH2O 19.7 μL。PCR循環條件為:95℃5 min;57.1℃1 min,72℃2 min,30個循環;最后72℃延伸10 min。取4 μL PCR產物進行1%瓊脂糖凝膠電泳檢測。
1.2.4 PCR產物的純化
將PCR產物進行1%瓊脂糖凝膠電泳后,切下擴增的目的條帶,利用OMEGA膠回收試劑盒回收PCR目的片段。
1.2.2 引物設計
1.2.5 熒光定量PCR
actin-a:5'ATCCTCCGTCTGGACTTGGC 3'
本文針對RBC的特點,考慮風險因素之間的影響,提出基于ANP和證據融合理論的風險評估模型,對RBC的風險等級進行評估。
利用Trizol reagent提取大豆食心蟲卵、幼蟲、蛹、成蟲四個時期和神經節、表皮、唾腺、中腸、卵巢、睪丸和脂肪體7個組織的總RNA,采用Fermentas公司M-MLV反轉錄酶合成cDNA。以cDNA為模板,利用熒光定量PCR對28SrDNA表達量進行檢測。熒光定量PCR按照TOYOBO公司SYBR?Green Realtime PCR Master Mix-Plus-程序進行。熒光定量PCR反應中所使用的基因及其引物如下:
actin-s:5'GGCGACATAGCACAGCTTCTC 3'
那時實行大集體制,生產隊按照家庭人口和掙得工分的多少,定期分給社員一些稻草、棉稈、松枝之類的東西用于做飯之用,但分配的那點柴草遠遠不能滿足炊用的需要,家家都感覺到柴不夠燒。棉稈、松枝還好燒一點,稻草一燒煙大灰多,做完一餐飯,是滿面火灰,兩眼通紅。所以,弄柴就成了我們這些孩子上學之余的主要任務,逢放學或放假,村子中10多個中小學生就都扛著鋤頭、背上箢子、帶上鐮刀浩浩蕩蕩地到山上找柴,遠山近坡能砍的都被我們砍光,能挖的都被我們挖完,山都變成了光禿禿的。
28S-a:5'ACGTTTGGTTCATCCCACAGC 3'
28S-s:5'CGGTAAAGCGAATGATTAGAGGC 3'
1.2.6 生物軟件
P1:AGGATTYCCYBAGTAGCKGCGAGCGAA;
1.2.7 載體構建
利用內切酶XbaⅠ和HindⅢ分別雙酶切具有雙向T7啟動子的載體L4440和28S rDNA目的片段,回收目標DNA片段,將純化產物連接,對重組子經菌液PCR鑒定,得到陽性克隆,命名為L4440-2828SrDNA。
2 結果與分析
2.1 大豆食心蟲28SrDNA基因克隆
從大豆食心蟲基因組模板獲得PCR擴增產物,引物1和引物2的擴增產物為1100 bp(見圖1a),引物3和引物4的擴增產物為1200 bp(見圖1b),并將PCR產物均進行測序,將兩段序列拼接成全長的大豆食心蟲28SrDNA序列,該序列長度為2127 bp。
在推進地下水超采治理試點工作中,始終牢牢把握五條基本原則:一是政府引導、全民行動,發揮好政府和群眾兩個積極性;二是規劃統領、科學治理,年度實施方案與中長期規劃有機銜接,集中連片規模實施,務求治理一片、見效一片、鞏固一片;三是創新機制、示范帶動,探索建立地下水超采治理的有效途徑,力求取得可示范、可復制、可推廣的經驗;四是因地制宜、積極穩妥,根據實際情況,科學確定治理模式和工程規模;五是競爭立項、績效考核,依據項目前期工作和壓采效果擇優實施,嚴格把關、嚴格獎懲。
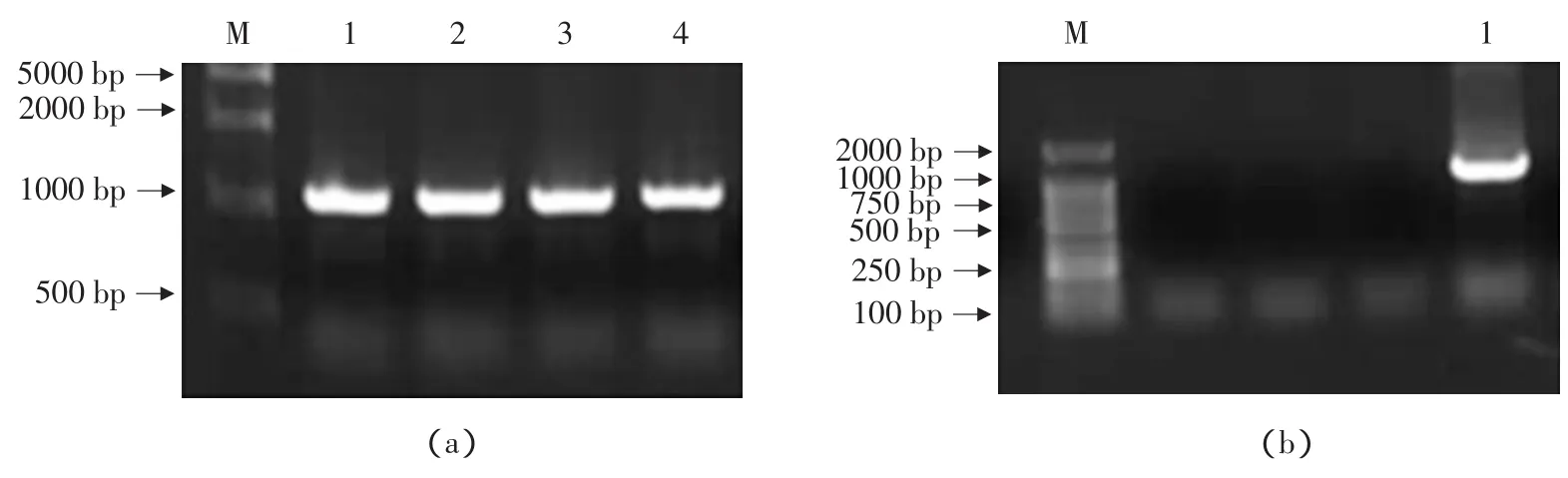
圖1 28SrDNA的PCR擴增結果Fig.1 PCR amplification results of 28SrDNA
將大豆食心蟲28SrDNA序列和其他昆蟲28SrDNA序列進行同源序列比較(見圖2),發現大豆食心蟲28SrDNA序列與蝎蛉(Anthocharis sara)、大蠟螟(Galleria mellonella)、粉蝶(Panorpa claripennis)、姬蜂(Poecilocryptus nigromaculatu)、家蠶(Bombyx mori)的28SrDNA同源性都在80%以上。采用MEGA4.0軟件利用相鄰連接法NJ(Neighborjoining method)構建系統進化樹,結果表明,大豆食心蟲的28SrDNA和鱗翅目家蠶的親緣關系較近,分布在同一分支,與膜翅目姬蜂親緣關系較遠,處于不同分支。
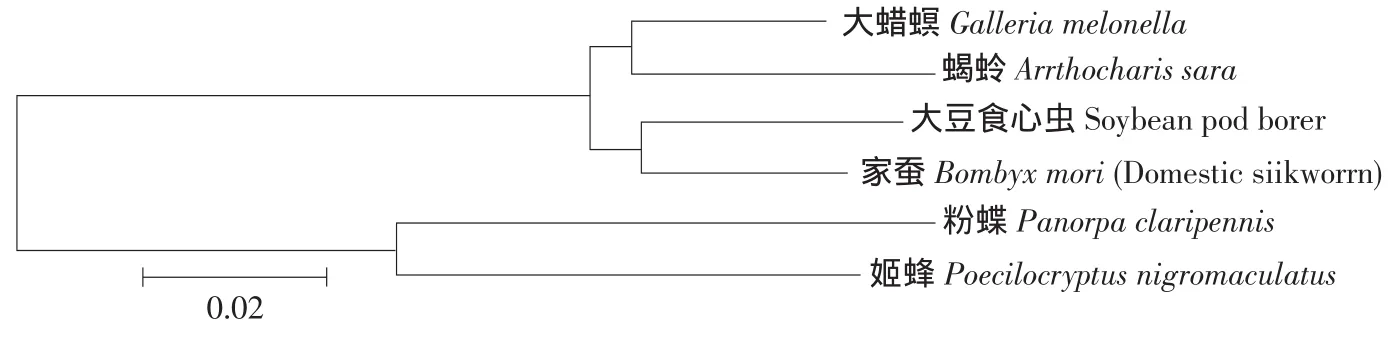
圖2 根據昆蟲28SrDNA序列利用相鄰連接(NJ)法構建的系統發生樹Fig.2 Phylogenetic analysis of the sequences of 28SrDNA from insect by neighbor joining method
2.2 大豆食心蟲28SrDNA基因的表達模式分析
為了確定大豆食心蟲28SrDNA基因在大豆食心蟲不同發育時期和大豆食心蟲幼蟲不同組織的表達情況,利用熒光定量PCR對28SrDNA mRNA的表達情況進行檢測,結果表明,28SrDNA的rRNA在卵、幼蟲、蛹、成蟲各發育階段均表達(見圖3),其中成蟲期(AD)28SrDNA的表達量最高,4齡幼蟲(N4)、蛹(PU)和1齡幼蟲(N1)的表達量較高,2齡幼蟲(N2)和3齡幼蟲(N3)表達量較低,卵中表達量最低。28SrDNA在神經節、表皮、唾腺、中腸、卵巢、睪丸和脂肪體7個組織中均有表達,其中脂肪體(FT)和睪丸(TE)的相對表達量最高,神經節(SY)的表達量較高,表皮(CU)、唾腺(SA)、中腸(MD)和卵巢(OV)相對表達量較低(見圖4)。
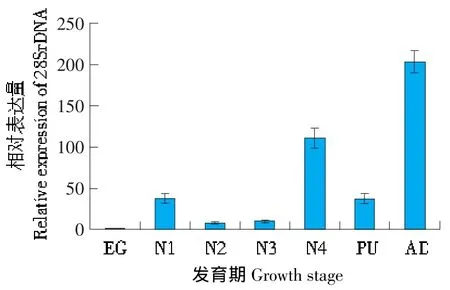
圖3 28SrDNA在大豆食心蟲不同發育階段的表達量Fig.3 Transcript level of 28SrDNA in different development stage
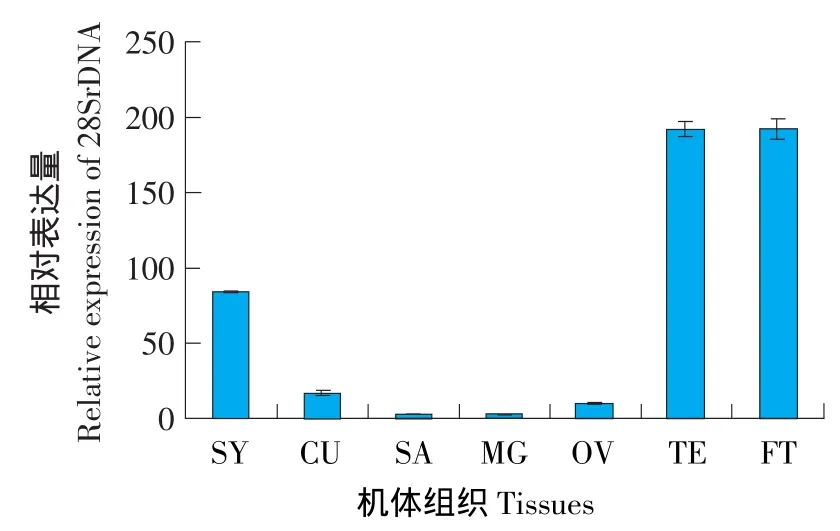
圖4 28SrDNA在大豆食心蟲不同組織的表達量Fig.4 Transcript level of 28SrDNA in different tissues of soybean pod borer
2.3 構建表達dsRNA的重組載體L4440-28SrDNA
利用DNAman軟件將大豆食心蟲28SrDNA和人28SrDNA進行序列比較,選擇同源性最低的大豆食心蟲28SrDNA序列區域作為目標片段。根據分析結果設計引物,通過PCR擴增得到418 bp的擴增產物(見圖5),純化后克隆到pMD18-T上。將質粒pMD18T-28S與載體L4440用XbaⅠ和HindⅢ雙酶切,回收目的片段,將純化產物連接,對重組子經菌液PCR鑒定,得到陽性克隆,命名為L4440-28SrDNA。重組質粒L4440-28SrDNA經XbaⅠ單酶切和XbaⅠ、HindⅢ雙酶切(見圖6),目的條帶均與預期結果相符,表明構建的重組載體L4440-28SrDNA正確。
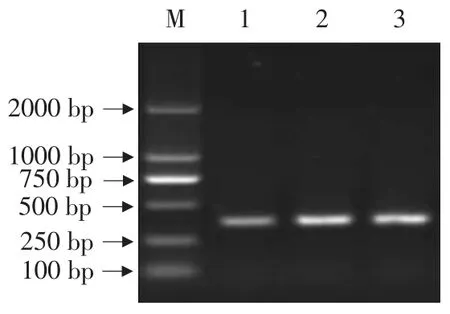
圖5 28SrDNA目的片段的PCR擴增產物Fig.5 PCR amplification result of 28SrDNA
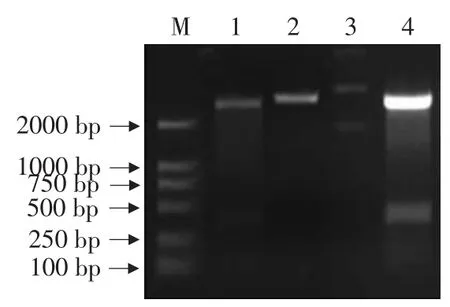
圖6 重組質粒L4440-28SrDNA的雙酶切鑒定Fig.6 Confirmation of recombinant plasmid L4440-28SrDNA by Xba I and Hind III
3 討論
RNA干擾是利用外源導入或轉錄載體在體內轉錄的雙鏈RNA介導受體細胞中的序列特異的同源性mRNA發生降解或翻譯受阻,引發受體細胞產生轉錄后基因沉默,使與此相應的內源靶基因的表達被阻抑,從而得到類似于“基因敲除”的基因阻抑效果[12]。Fire等在線蟲發現RNA干擾現象后,在真菌、植物、線蟲、昆蟲、斑馬魚、小鼠等大多數真核生物也發現RNAi現象,研究表明RNAi是生物進化過程中保留下來的一種抵抗微生物侵入、轉座子擴展和對基因表達進行轉錄后調控的強大機制。由于RNAi沉默基因的高效性和特異性,將昆蟲關鍵生長基因dsRNA導入昆蟲體內細胞,dsRNA介導受體細胞內同源性mRNA發生降解,使昆蟲體內與dsRNA相對應的靶基因表達被抑制,可以直接分析基因的功能。目前的主要問題是尋找一個向目標昆蟲體內導入dsRNA簡單而且穩定的方法。Timmons等發現給線蟲喂食表達dsRNA的大腸桿菌或將線蟲浸泡在dsRNA溶液中均能誘導線蟲體內靶基因的沉默,表明dsRNA可以直接從環境(體內腸道和體表)進入到昆蟲體內[13]。
目前,國內外尚無大豆食心蟲RNA干擾的研究報道。本研究以大豆食心蟲幼蟲作為研究對象,利用同源克隆法克隆大豆食心蟲28SrDNA全長序列。對大豆食心蟲28SrDNA不同發育階段和不同組織的表達量進行分析,結果表明,大豆食心蟲28SrDNA在大豆食心蟲的不同發育階段的不同組織均有表達,在大豆食心蟲生長發育階段起關鍵作用。
本研究將28SrDNA的目的片段克隆到L4440載體上,該載體具有雙T7強啟動子,對目標基因片段的克隆不要求方向性,目的片段以何種方向連入,利用該載體的雙T7強啟動子均可以得到對應于目標基因的dsRNA。構建完的載體可轉入E.coli HT115(DE3)中,該菌株含有T7RNA聚合酶基因,在T7強啟動子的控制下進行該基因的轉錄,產生T7RNA聚合酶。而且該菌株還是一個Rnc基因突變菌株,不能編碼降解dsRNA的內切酶RnaseⅢ的基因,這樣在IPTG的誘導下,在E.coli HT115菌株中很容易產生大量的目的片段的dsRNA。因此該載體的成功構建,為利用喂食方法研究大豆食心蟲28SrDNA基因的功能奠定基礎,有利于進一步探索利用植物介導的RNA技術防治大豆食心蟲。
[1] 劉洋,王繼安,趙奎軍.大豆抗食心蟲性遺傳研究[J].東北農業大學學報,2005,36(2):138-141.
[2] Fire A,Xu S Q,Montgomery M K,et al.Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J].Nature,1998,391:806-811.
[3] Baum J A,Bogaert T,Clinton W,et al.Control of coleopteran insect pests through RNA interference[J].Nature Biotechnology,2007,25:1322-1326.
[4] Whyard S,Singh A D,Wong S.Ingested double-stranded RNAs can act as species-specific insecticides[J]Insect Biochemistry and Molecular Biology,2009,39:824-832.
[5] Yadav B C,Veluthambi K,Subramaniam K.Host-generated double stranded RNA induces RNAi in plant-parasitic nematodes and protects the host from infection[J].Molecular&Biochemical Parasitology,2006,148:219-222.
[6] Steeves R M,Todd T C,Essig J S.Transgenic soybeans expressing siRNAs specific to a major sperm protein gene suppress Heterodera glycines reproduction[J].Functional Plant Biology,2006,33:991-999.
[7] Price D R,Gatehouse J A.RNAi-mediated crop protection against insects[J].Trends in Biotechnology,2008,26:393-400.
[8] MaoY B,Cai W J,Wang J W.Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol[J].Nature Biotechnology,2007,25:1307-1313.
[9] Baum J A,Bogaert T,Clinton W.Control of coleopteran insect pests through RNA interference[J].Nature Biotechnology,2007,25:1322-1332.
[10] 劉望夷.核糖體的生物功能、自我剪切與自我復制[J].生物化學與生物物理進展,1991,18(1):1-5.
[11] 潘敏慧,萬永承,魯成.核糖體RNA研究進展[J].蠶學通訊,2001,21(3):10-14.
[12] Bischoff V,Vignal C,Duvic B,et al.Down regulation of the Drosophila immune response by peptidoglycan-recognition proteins SC1 and SC2[J].PLoS Pathogens,2006(2):139-147.
[13] Timmons L,Court D L,Fire A.Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans[J].Gene,2001,263:103-112.
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
