白僵菌培養基繼代培養對蛋白酶(cdep1)、幾丁質酶(chit1)基因表達的影響
2012-02-28 07:47:56孫召朋張正坤徐文靜汪洋洲潘洪玉李啟云
植物保護 2012年2期
孫召朋, 張正坤, 徐文靜, 汪洋洲, 潘洪玉, 李啟云*
(1.吉林大學,長春 130062; 2.吉林省農業科學院,長春 130033)
白僵菌(Beauveria bassiana)是目前國內生物防治中應用最廣泛的昆蟲病原真菌[1],具有防治害蟲效果好,不傷害天敵,對人、畜、植物無毒害,容易大量生產,無殘毒,害蟲不易產生抗性等優點[2],在玉米螟和松毛蟲等的防治中已取得很好的防效[3]。研究表明,在穿透寄主體壁的過程中,球孢白僵菌會產生多種降解酶。在穿透昆蟲體壁的早期,昆蟲病原真菌高水平表達蛋白酶,降解昆蟲的體表蛋白[4]。同時,也表達用于降解昆蟲體壁的主要組成成分幾丁質的幾丁質酶[5]。球孢白僵菌在防治應用中菌種資源主要來源于自然直接篩選的菌株,生產的菌株“退化”是防效不穩定的重要原因之一。高毒力菌株經多代培養后毒力退化的現象已被認識[6],但原因不是十分清楚。本研究對兩株田間采集分離的野生白僵菌菌株進行了培養基繼代培養,通過對亞洲玉米螟的室內毒力測定,評價了繼代培養對白僵菌毒力的影響。同時,明確了每一世代白僵菌蛋白酶和幾丁質酶基因的表達水平,分析了繼代培養對白僵菌毒力及其蛋白酶和幾丁質酶基因的影響。
1 材料與方法
1.1 試驗材料
供試昆蟲:亞洲玉米螟公主嶺種群[Ostrinia f ur nacalis(Guenée)]2齡幼蟲,由吉林省農業科學院植物保護研究所汪洋洲博士提供。
供試菌株:球孢白僵菌D1-5和D6-2分離自德惠市玉米田的玉米螟僵蟲蟲體,由本實驗室沙土管保存于-80℃超低溫冰箱(Ther mo For ma)。
1.2 主要試劑
限制性核酸內切酶Eco RⅠ、Hin dⅢ購自MBI Fer mentas,Trizol reagent購自上海生工,瓊脂糖為Bio-Rad公司產品,總RNA抽提試劑盒為大連寶生物工程有限公司(Ta Ka Ra)產品。所用無水乙醇、異丙醇、氯仿等有機溶劑為中國醫藥(集團)上海化學試劑公司產品,均為分析純。
1.3 室內毒力測定
1.3.1 白僵菌供試菌株的繼代培養
將供試菌株在無菌條件下從沙土管接種在SDAY培養基(質量濃度4%)葡萄糖,1%蛋白胨,2%酵母膏,p H7.0),為F1代,培養20 d左右后刮取孢子,將孢子制備成1×107個/mL濃度的孢子懸液,涂布于SDAY培養基,共進行F2~F5代培養,每個菌株各世代培養10皿,收集每個世代孢子,均配制成1×107孢子/mL濃度的孢子懸浮液供試。
1.3.2 萌發率的測定
取相同量菌懸液接種于SDAY培養基中(終濃度為1×105孢子/mL),26℃培養16 h,測其萌發率,每個菌株重復3次。
1.3.3 室內生物測定方法
將室內飼養、齡期一致的2齡玉米螟幼蟲浸入1×107孢子/mL的孢子懸液中5~10 s后,用濾紙吸去多余藥液,將試蟲轉移到正常條件下飼養。每處理4次重復,每重復浸蟲20頭,并設無菌水處理作為對照。在黑暗環境下培養10 d,每天記錄被白僵菌感染的試蟲數。
1.3.4 數據統計與分析方法
根據調查數據,計算各處理的校正死亡率,公式如下:
死亡率(P1)=死亡蟲數(K)/處理總蟲數(N)×100%;
校正死亡率(P2)=[處理死亡率(Pt)-空白對照死亡率(P0)]/[1-空白對照死亡率(P0)];
利用DPS分析軟件進行統計分析數據。
1.4 半定量RT-PCR檢測繼代培養對白僵菌蛋白酶、幾丁質酶基因表達的影響
1.4.2 樣品收集及總RNA提取
從培養供試菌株的PDA培養基上刮取100 mg分生孢子,采用Trizol法提取總RNA。利用紫外分光光度計測定各處理中總RNA的濃度,并保證A260/A280比值在 1.8~2.0之間,以確保總 RNA純度。
1.4.2 半定量 RT-PCR[7-11]
利用紫外分光光度計測定樣品RNA濃度,每個處理上樣量為50 ng總RNA,利用蛋白酶(cdep1)(AY040532.1)、幾丁質酶(chit1)(AY145-440.1)及管家基因18S r DNA(GQ302680.1)特異性引物進行c DNA第1鏈合成。以各基因c DNA第1鏈為模板進行目的片段PCR,反應中調整反應循環數,以10、15、20個循環每隔5個循環進行一次反應,PCR反應條件為以目的基因條帶指數增長期循環數為準確定基因表達量的差異。各基因PCR反應體系為:2μL c DNA產物,0.5μL Taq DNA聚合酶,目的基因(或18Sr DNA基因)上下游引物各1μL,d NTPs 1μL,10×buffer 5μL,Mg Cl2(25 mmol/L)3μL,終體積50μL。反應程序如下:94℃2 min,94℃50 s,58℃(cdep1)/52 ℃ (chit1)/58 ℃(18S r DNA)50 s,72 ℃ 1 min (10、15、20、25個循環),72℃10 min。4℃保存。
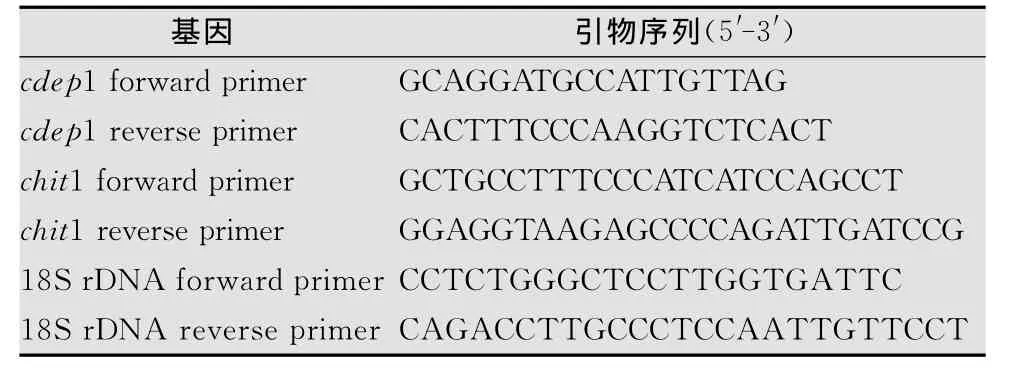
表1 待測基因所用引物序列
根據上述PCR反應結果確定各基因擴增指數期循環數,以供試菌株不同世代各基因c DNA為模板,進行PCR擴增。瓊脂糖凝膠電泳膠濃度1.5%。
2 結果與分析
2.1 繼代培養對白僵菌孢子萌發率的影響
兩菌株在SDAY培養基中經16 h培養后,其萌發率如圖1所示。兩個菌株的萌發率變化規律基本相似,F2代萌發率明顯高于F1代,分別達到77%和70%,在隨后的繼代培養中,F3、F4、F5代萌發率依次降低,L1-1的F5代萌發率僅為2%。
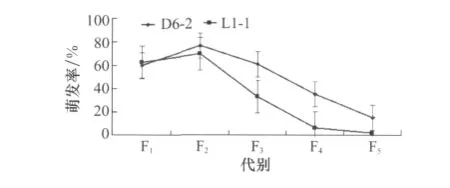
圖1 供試白僵菌菌株不同世代萌發率測定(p=0.05)
2.2 繼代培養對白僵菌殺玉米螟毒力的影響
白僵菌兩菌株不同世代對亞洲玉米螟2齡幼蟲的毒力測定結果見表2和表3。菌株L1-1 F1代孢子對亞洲玉米螟的致死中時(LT50)和致死90%時間(LT90)分別為186.63 h和284.16 h,而F2代孢子分別為180.61 h和209.61 h,明顯低于F 1代,而F3~F5代孢子的LT50和LT90逐漸增加,明顯高于F2代和F1代,說明其毒力在不斷下降。D6-2菌株F1代孢子對亞洲玉米螟的LT50和LT90分別為170.09 h和190.21 h,而 F2代分別為137.70 h和165.84 h,明顯低于F1代,而F3~F5代孢子的LT50和LT90逐漸增加,明顯高于F2代和F1代,說明隨著繼代次數的增加其毒力也在不斷下降。
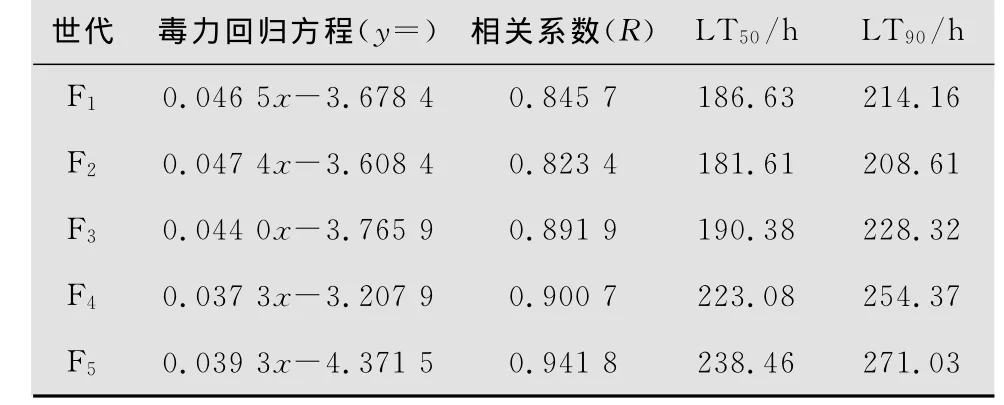
表2 白僵菌L1-1菌株不同世代對亞洲玉米螟2齡幼蟲的毒力(10 7孢子/mL)
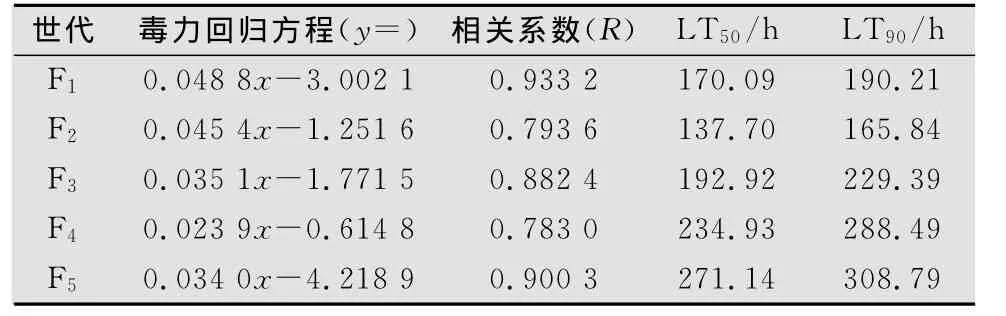
表3 白僵菌D6-2菌株不同世代對亞洲玉米螟2齡幼蟲的毒力(10 7孢子/mL)
2.3 繼代培養對白僵菌毒力相關基因表達的影響
2.3.1 RT-PCR 反應循環數的確定
利用供試菌株目的基因和管家基因c DNA第1鏈為模板,進行目的基因半定量RT-PCR反應循環數的確定。從圖2中可以看出,18S r DNA基因和chit 1基因在15~20個循環時條帶差異明顯,而cdep 1基因在10~15個循環時條帶差異明顯,因此,以18個循環為18S r DNA基因和chit 1基因的檢測循環數,以13個循環作為cdep1基因的檢測循環數。
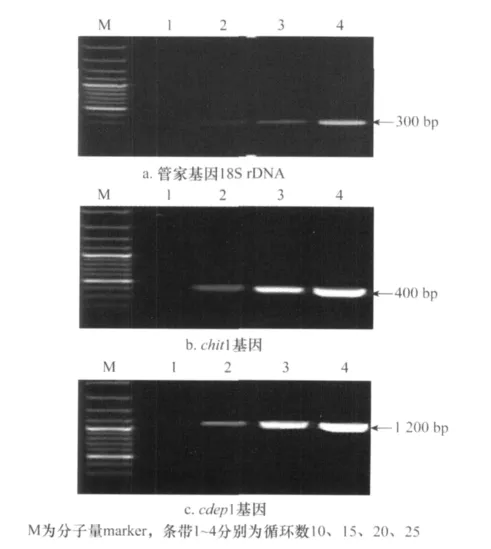
圖2 管家基因及毒力相關基因半定量RT-PCR循環數的確定
2.3.2 目的基因的半定量RT-PCR檢測
為了明確繼代培養對球孢白僵菌毒力相關酶基因表達情況的影響,以18Sr DNA基因為內參,利用半定量RT-PCR對cdep 1基因和chit 1基因的表達情況進行了檢測。從圖3可以看出,在內參基因穩定表達,也就是總RNA量相同的情況下,兩個供試菌株L1-1和D6-2的cdep 1基因和chit1基因在F 1和F2代表達穩定,隨著繼代次數的增多,供試菌株兩個基因的PCR條帶明顯變弱,說明其表達量在不斷下降,這與對亞洲玉米螟毒力測試的結果一致。
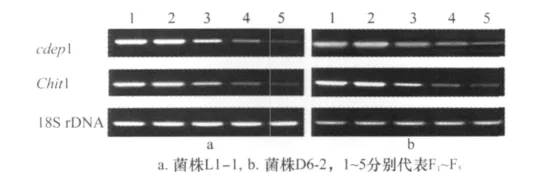
圖3 繼代培養對球孢白僵菌毒力相關酶基因cdep1和chit1基因mRNA表達的影響
3 討論
毒力測定結果表明,兩株白僵菌供試菌株的F2代比F1代對亞洲玉米螟2齡幼蟲的毒力有所增強,而F3~F5代對玉米螟的毒力呈下降趨勢。球孢白僵菌通過分泌蛋白酶、幾丁質酶和脂酶來降解昆蟲體壁,其中幾丁質酶和蛋白酶具有重要作用[12-14]。本研究利用半定量RT-PCR方法對兩株白僵菌中兩種毒力相關酶基因在不同世代的表達情況進行了測定,發現隨著繼代次數的增多,供試菌株兩個毒力相關酶基因的表達量均呈下降趨勢,與對亞洲玉米螟毒力測定結果一致。出現這種現象可能是由于F1代直接接種于沙土管,菌株并沒有表現出很高的萌發率和毒力,在F1活化之后,F2代達到其毒力最大值,F3~F5依次減弱。
在白僵菌繼代培養過程中,隨著繼代次數的增多,會有多個生物性狀發生改變。P.Rajanikant h等[15]報道繼代培養對產孢量和毒力有很大的影響。唐曉慶等[6]報道了繼代培養中菌落都會發生變化,并提出了菌落局變發生的幾種可能的遺傳機制。唐曉慶等人[16]報道了繼代培養對抗旱力的影響,結果顯示繼代培養能增強菌株的抗旱力,但菌株生活力減弱。樊美珍等[17]通過研究證明,以SDAY為基礎培養基的不同C/N比、不同p H培養基中,均未發現菌落局變及菌種退化,SDAY最適于白僵菌生長。本試驗中采用SDAY為培養基研究供試白僵菌菌株萌發率、毒力及其相關酶基因的表達,結果表明這些指標均發生了變化,并且表現出相似的變化規律。F1與F2代各項指標基本相似或者略低于F 2代,從F2代開始到F5代,萌發率、毒力及其相關酶基因的表達呈現逐漸降低的趨勢,這和唐曉慶等人所報道的繼代培養中產生不同類型的分離株,這些分離株多表現為產孢量下降、毒力降低等與生產性狀退化類似的變異現象基本相符[18]。其可能的原因是由于F1剛從沙土管中接出,菌株新陳代謝能力下降,經過相當于活化階段的F1代后,在F2代各種指標都達到最大值,從F2代到F5代的繼代培養,菌株毒力相關酶基因表達量逐漸降低,從而導致了毒力的下降。Ansari等[19]對另一廣泛應用的生防真菌綠僵菌(Metar hiziu m anisopliae)的3株商業化菌株進行了12代的繼代培養,發現其毒力并未發生變化,并將其作為綠僵菌高效菌株的篩選標準。生防真菌的毒力在繼代培養過程中可能依賴其自身的遺傳背景,本研究中只對兩株白僵菌菌株進行了繼代培養對毒力及毒力相關因素的影響,是否在白僵菌中存在經繼代培養而毒力不發生變化的菌株,還需要進一步研究。
由于白僵菌對靶標昆蟲的致病力由多種因素造成[6],本研究只是從蛋白酶和幾丁質酶基因的表達方面進行了研究,關于其他影響因素如萌發率、毒素代謝水平等還需進一步研究。同時,在菌種使用過程中,定期進行蟲體復壯,可以恢復菌株毒力這一問題上存在爭論[20],本試驗只對培養基繼代培養白僵菌毒力退化情況進行了測定,對于通過蟲體進行菌株復壯并未研究,后者可作為以后研究的新方向。
[1] 林海萍,韓正敏,張昕,等.球孢白僵菌研究現狀及提高其殺蟲效果展望[J].浙江林學院學報,2006,23(5):575-580.
[2] 馮明光.胞外蛋白酶和脂酶活性作為球孢白僵菌毒力指標的可靠性分析[J].微生物學報,1998,38(6):461-467.
[3] 王清海,萬平平,黃玉杰,等.蟲生真菌在害蟲生物防治中的應用研究[J].山東科學,2005,18(4):37-41.
[4] Goettel M S,Hajek A E,Siegel J P,et al.Safety of f ungal biocontrol agents[M]∥Butt T M,Jackon C,Magan N.Fungi as biocontr ol agents,CAB Inter national,2001:347-375.
[5] Char nley A K,St.Leager R J.The role of cuticle-degrading enzy mes in f ungal pat hogenesis in insects[M]∥Cole E T,Hoch H C.Fungal spore disease initiation in plants and animals New York,London:Plenu m Press,1991:267-287.
[6] 唐曉慶,樊美珍,李增智.球孢白僵菌繼代培養中菌落局變現象及環境影響因素的研究[J].真菌學報,1996,15(3):45-53.
[7] 周林福,陳離偉,姜云水,等.熒光定量PCR與半定量PCR檢測HBV DNA的對比分析[J].中國病理生理雜志,2005,21(5):837,848.
[8] 邊杉,王穎,于濤,等.聚丙烯酰胺凝膠銀染技術在半定量RT-PCR中的應用[J].中國藥科大學學報,2004,35(2):178-182.
[9] 石艷麗,郭學平,王鳳山,等.半定量RT-PCR法測定鏈球菌透明質酸合酶 mRNA 的水平[J].食品與藥品,2005,7(2 A):22-24.
[10]徐春蘭,汪以真.半定量RT-PCR法分析豬肌肉組織細胞谷胱甘肽過氧化物酶mRNA表達水平[J].中國獸藥雜志,2005,39(8):3-6.
[11]陳昕,王保莉,曲東,等.小麥硫轉運蛋白基因半定量RT-PCR檢測方法的建立[J].西北植物學報,2006,26(2):309-313.
[12]范艷華.球孢白僵菌降解寄主體壁的幾丁質酶和蛋白酶的分子改良[D].重慶:西南大學,2006.
[13]Fang Weiguo,Leng Bo,Xiao Yuehua,et al.Cloning of Beauveria bassiana chitinase gene bbchit1 and its application to i mpr ove f ungal strain vir ulence[J].Applied and Envir on mental Microbiology,2005,71(1):363-370.
[14]Yanhua Fan,Weiguo Fang,Yuehua Xiao,et al.Directed evol ution for increased chitinase activity[J].Appl Micr obiol Biotechnol,2007,76:135-139.
[15]Rajanikant h P,Subbarat na m G V,Raha man S J,et al.Effectof frequency of subculturing of different isolates of Beauveria bassiana Vuillemin on their biological pr operties[J].Bio-resource Manage ment,2011,2(1):60-65.
[16]唐曉慶,黃勃,李增智.繼代培養對球孢白僵菌抗旱力的影響[J].安徽農業大學學報,1996,23(3):289-292.
[17]樊美珍,李增智,唐曉慶.白僵菌菌種退化及其控制[J].安徽農業大學學報,1996,23(3):239-245.
[18]唐曉慶,唐燕平,李增智.球孢白僵菌菌種退化及其對馬尾松毛蟲防治的影響[J].安徽農業大學學報,1996,23(3):246-253.
[19]Ansari M A,Butt T M.Effects of successive subculturing on stability,vir ulence,conidial yield,ger mination and shelf-life of ento mopat hogenic f ungi[J].Jour nal of Applied Micr obiology,2011,110:1460-1469.
[20]Hall R A.Effectof repeated subcult uring on agar and passaging t hr ough an insect hoston pat hogenicit y,mor phology,and growt h-rate of Verticilliu m lecanii[J].Jour nal of Invertebrate Pat hology,1980,36(2):216-222.
