三個小麥農家品種的苗期抗白粉病遺傳分析
2012-02-28 07:47:54趙小華許紅星李秀全安調過
植物保護 2012年2期
趙小華, 許紅星, 李秀全, 安調過*
(1.中國科學院遺傳與發育生物學研究所農業資源研究中心,石家莊 050021;2.中國農業科學院作物科學研究所,北京 100081;3.中國科學院研究生院,北京 100049)
小麥白粉病是由布氏白粉菌小麥專化型(Blumeria gr a minis f.sp.tritici,Bgt)引起的真菌病害,對我國小麥安全生產造成嚴重威脅。培育抗病品種是最為經濟、安全、有效的防治措施[1]。現代農業造成的抗源及抗病基因的單一化,加速了對不斷變異的白粉菌小種的定向選擇作用,促進毒性新小種不斷出現,使得小麥生產品種的抗性不斷喪失。因此有必要挖掘新的抗源材料,提高抗病基因的多樣性,從而有效控制小麥白粉病。
目前國際上正式命名的小麥抗白粉病主效基因有57個,分布在40個基因位點上(Pm1~Pm43),這些基因來源于普通小麥、小麥近緣屬和小麥近緣種,其中來自普通小麥的抗病基因有Pm4c[2]、Pm38[3]和Pm39[4]等27個,其余的來自于小麥近緣屬(黑麥、山羊草、簇毛麥以及中間偃麥草等)和小麥近緣種(栽培一粒小麥、野生一粒小麥、波斯小麥等)。已經報道的小麥抗白粉病基因中,Pm5e、ml xbd和Pm24分別來源于我國的小麥農家品種‘復壯30’[5]、‘小白冬麥’[6]和‘齒牙糙’[7]。小麥農家品種是經過長期自然和人工選擇的產物,具有豐富的遺傳多樣性。對其抗病基因進行研究,并進行合理的開發和利用,將有助于改善抗病基因的單一化,增加抗源的多樣性。
本實驗室對258份國內小麥農家品種苗期接種國內北方麥區流行的白粉菌E09菌株和石家莊地區的混合白粉病原菌,篩選出‘矮稈芒麥’(ZM04132)、‘紅頭麥’(京2651)和‘大紅頭’(S761)表現免疫至高抗,繼續對這3份農家品種接種白粉菌E03、E05、E18、E20和E23菌株,結果均表現為抗病。為進一步研究它們的抗白粉病遺傳特點,本研究對這3份品種進行苗期抗性的遺傳分析,為其在抗病育種中的利用提供依據。
1 材料與方法
1.1 材料
供試小麥農家品種‘矮稈芒麥’、‘紅頭麥’和‘大紅頭’來自于國家種質資源庫,并由前期白粉病抗性鑒定篩選而來;作為感病對照的普通小麥品種‘銘賢169’由本實驗室保存。小麥白粉菌E09菌株由中國農業科學院植物保護研究所提供。
1.2 抗性鑒定
所有小麥材料均播種于72孔的穴盤中,每個穴盤隨機種植‘銘賢169’作為感病對照。所有小麥材料播種后置于室溫下培養。待苗生長至一葉期時進行接種,置于黑暗條件下保濕培養24 h后,放在溫室中培養,L∥D=14 h∥10 h,晝夜溫度為22℃/18℃。接種后10 d,待‘銘賢169’充分發病,調查記載發病情況。抗性調查時,反應型按照“0~4”六級記載[8];0型為免疫,0;型為近免疫,1~4型分別為高抗、中抗、中感和高感。
1.3 統計分析
根據調查記載的情況,將0~2型劃分為抗病類型,3~4型劃分為感病類型,計算雜交F2代群體的抗病單株與感病單株的比值,并經卡方測驗分析其符合度。參照何家泌[9]小麥抗病基因分離理論模式,結合F1植株的抗病性、F2群體的抗感單株分離比例,對鑒定結果進行分析。
2 結果與分析
2.1 小麥農家品種‘矮稈芒麥’抗性遺傳分析
‘矮稈芒麥’對白粉菌E09菌株的反應型為0~1型,‘銘賢169’對E09的反應型為4型,正交和反交的F1代植株對E09的反應型均為4型,表明‘矮稈芒麥’對E09的抗性主要由隱性核基因控制。在161株‘矮稈芒麥’ב銘賢169’組合構成的F2分離群體中,有47株表現抗病、114株表現感病。抗病株中反應型為0型的有18株、反應型為1型的有13株、反應型為2型的有16株;感病株中反應型為3型的有55株、反應型為4型的有59株;抗感分離比符合1∶3(χ2=1.29,p=0.26,表1)。遺傳分析結果表明,‘矮稈芒麥’對E09的抗性由1對隱性基因控制[9]。
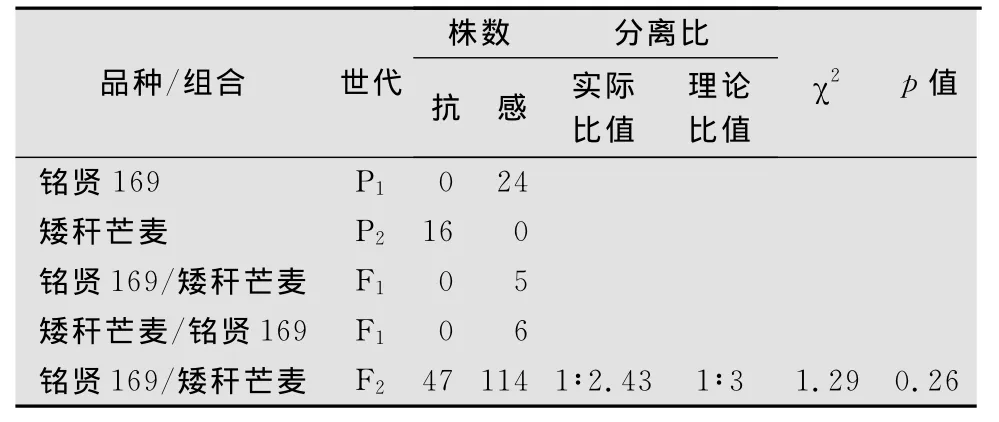
表1 ‘矮稈芒麥’與‘銘賢169’雜交各世代對白粉菌E09菌株的抗性表現
2.2 小麥農家品種‘紅頭麥’抗性遺傳分析
遺傳分析與2.1類似,‘紅頭麥’對白粉菌E09菌株的反應型為0型,‘銘賢169’對E09的反應型為4型,正交和反交的F1代植株對E09的反應型均為4型,表明‘紅頭麥’對E09的抗性主要由隱性核基因控制。在156株‘紅頭麥’ב銘賢169’組合構成的F2分離群體中,有42株表現抗病、114株表現感病。抗病株中反應型為0型的植株有5株、反應型為1型的有6株、反應型為2型的有31株;感病株中反應型為3型的有45株、反應型為4型的有69株;抗感分離比符合1∶3(χ2=0.21,p=0.64,表2)。遺傳分析結果表明,‘紅頭麥’對E09的抗性由1對隱性基因控制。
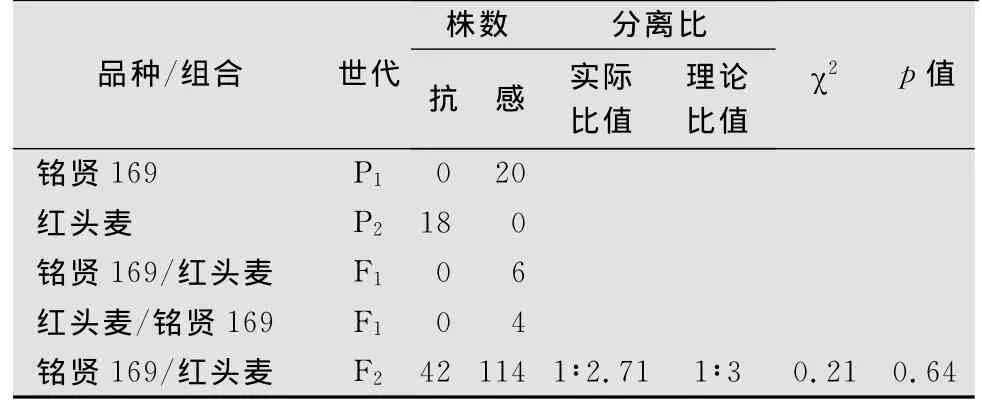
表2 ‘紅頭麥’與‘銘賢169’雜交各世代對白粉菌E09菌株的抗性表現
2.3 小麥農家品種‘大紅頭’抗性遺傳分析
遺傳分析與前面兩者類似,‘大紅頭’對白粉菌E09菌株的反應型為0型,‘銘賢169’對E09的反應型為4型,正交和反交的F1代植株對E09的反應型均為4型,表明‘大紅頭’對E09的抗性主要由隱性核基因控制。在157株‘紅頭麥’ב銘賢169’組合構成的F2分離群體中,有43株表現抗病、114株表現感病。抗病株中反應型為0型的植株有20株、反應型為1型的有13株、反應型為2型的有10株;感病株中反應型為3型的有80株、反應型為4型的有34株;抗感分離比符合1∶3(χ2=0.36,p=0.55,表3)。遺傳分析結果表明,‘大紅頭’對E09的抗性由1對隱性基因控制。
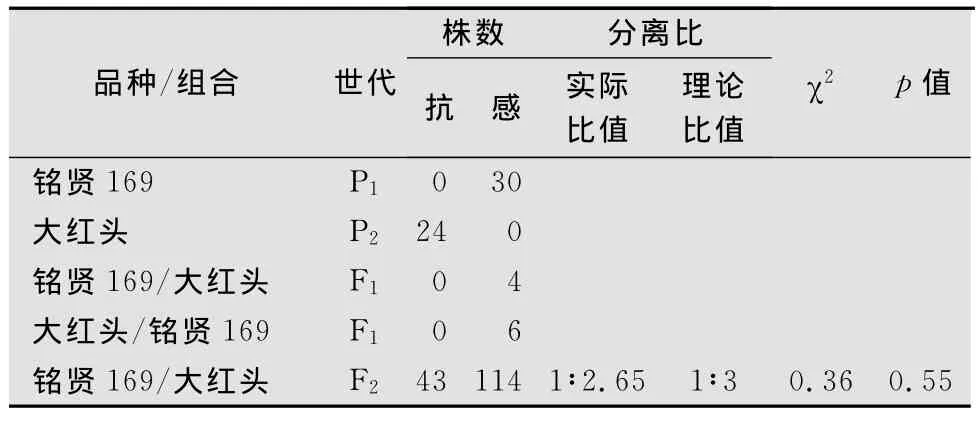
表3 ‘大紅頭’與‘銘賢169’雜交各世代對白粉菌E09菌株的抗性表現
3 討論
小麥農家品種‘矮稈芒麥’、‘紅頭麥’和‘大紅頭’對石家莊本地區的混合白粉病菌表現出良好的抗性。本研究利用白粉菌E09菌株,對這3份農家品種進行了抗性遺傳分析。結果表明,這3份農家品種對E09的抗性均由1對隱性基因控制。該結果有助于這3份農家品種進一步的分子標記及定位研究,為其作為抗源在小麥抗白粉病育種中的應用奠定了基礎。
對小麥農家品種中的白粉病抗性基因進行遺傳分析,前人也有一些報道[10-12]。其中胡衛國等對陜西小麥農家品種‘矮稈芒麥’和‘大紅頭’進行白粉病抗性遺傳分析,結果表明,‘矮稈芒麥’含有3對隱性抗白粉病基因,‘大紅頭’含有2對顯性互補抗白粉病基因[10],與本研究的結果并不一致。胡衛國等采用的白粉病菌株為關中4號,本研究采用的白粉菌為E09菌株,不同的菌株有不同的毒性(無毒性)基因,品種中與之相對應的抗性基因也不同,因此有不同的抗性表現;另外,小麥農家品種是我國農民在長期的生產實踐中選擇、經品種資源科學家征集、整理而來的品種,往往多個材料具有相同的名字,不同來源和材料本身不同的情況時常發生,胡衛國等用到的小麥農家品種‘矮稈芒麥’、‘大紅頭’與本研究中的‘矮稈芒麥’、‘大紅頭’同名但也有可能來源不同。總之,農家品種是改良小麥的一個重要基因庫。由于農家品種本身普遍存在的異質性,因此,對農家品種的研究,首先要在保證材料純化的基礎上,進一步鑒定發掘其攜帶的有益基因。本研究涉及的3份農家品種都是在這樣的基礎上進行抗病性鑒定和遺傳分析的。
目前已正式命名的小麥抗白粉病基因中,大多數表現為顯性遺傳,少數表現為隱性遺傳,包括Pm5a、Pm5b、Pm5c、Pm5d、Pm5e、Pm9、Pm26 和Pm42[13-14]。本研究中發現的3份小麥農家品種‘矮稈芒麥’、‘紅頭麥’和‘大紅頭’所攜帶的抗性基因均為隱性基因。Pm5a的載體品種‘Hope’對白粉菌E03、E05、E09、E20和 E23菌株均表現為感病[15],Pm5b的載體品種‘Kor moran’對E09表現出感病[16],Pm5c的載體品種‘Kolandi’對 E05、E09表現出感病[16],Pm5e的載體品種‘復壯30’對 E18、E23表現出感病[17],而本研究中3份農家品種‘矮稈芒麥’、‘紅頭麥’和‘大紅頭’對 E03、E05、E09、E18、E20和E23這些菌株均表現抗病,可以推知,這3份農家品種所攜帶的白粉病抗性基因應該不同于Pm5a、Pm5b、Pm5c、Pm5e。另外,兩個抗白粉病隱性基因Pm26[18]和 Pm42[14]都來源于野生二粒小麥,而本研究中的白粉病抗性基因均來自于農家品種,推測這3份農家品種所攜帶的白粉病抗性基因可能不同于Pm26和Pm42。至于3份農家品種中含有的抗白粉病基因與Pm5d和Pm9是否相同,還有待于進一步的分析。而3份農家品種所攜帶的抗白粉病基因之間的相互關系,也有待于進一步的等位性分析。
[1] 邵振潤,劉萬才.我國小麥白粉病的發生現狀與治理對策[J].中國農學通報,1996,12(6):21-23.
[2] Hao Y,Liu A,Wang Y,et al.Pm23:a new allele of Pm4 located on chromosome 2 AL in wheat[J].Theor Appl Genet,2008,117:1205-1212.
[3] Spiel meyer W,Singh R P,Mc Fadden H,et al.Fine scale genetic and physical mapping using interstitial deletion mutants of Lr34/Yr 18:a disease resistance locus effective against multiple pathogens in wheat[J].Theor Appl Genet,2008,116:481-490.
[4] Lillemo M,Asalf B,Singh R P,et al.The adult plant r ust resistance loci Lr 34/Yr 18 and Lr46/Yr 29 are i mportant deter minants of partial resistance to powdery mildew in bread wheat line Saar[J].Theor Appl Genet,2008,116:1155-1166.
[5] Huang X Q,Wang L X,Xu M X,et al.Microsatellite mapping of the powder y mildew resistance gene Pm5e in co mmon wheat(Triticu m aestivu m L.)[J].Theor Appl Genet,2003,106:858-865.
[6] Huang X Q,Hsam S L K,Zeller F J.Chromosomal location of t wo novel genes for resistance to powder y mildew in Chinese landraces(Triticu m aestivu m L.em.Thell.)[J].J Genet Breed,2000,54:311-317.
[7] Huang X Q,Hsam S L K,Zeller F J.Chro moso mal location of genes for resistance to powdery mildew in common wheat(Triticu m aestivu m L.em.Thell.)4.Gene Pm24 in Chinese landrace Chiyacao[J].Theor Appl Genet,1997,95:950-953.
[8] 盛寶欽.用反應型記載小麥苗期白粉病[J].植物保護,1988,14(1):49.
[9] 何家泌.植物抗病遺傳學[M].北京:中國農業出版社,1994.
[10]胡衛國,王亞娟,王長有,等.陜西小麥地方品種白粉病抗性的遺傳分析[J].麥類作物學報,2007,27(2):341-344.
[11]翟雯雯,段霞瑜,周益林,等.我國小麥地方品種螞蚱麥、小白冬麥、游白蘭、紅卷芒麥抗白粉病性遺傳分析[J].植物保護,2008,34(1):37-40.
[12]王錫鋒,張忠山,劉紅彥,等.河南農家小麥品種資源抗、慢白粉病性鑒定[J].河南農業大學學報,1996,30(2):160-164.
[13]解超杰,楊作民,孫其信.小麥抗白粉病基因[J].西北植物學報,2003,23(5):822-829.
[14]Hua W,Liu Z J,Zhu J,et al.Identification and genetic mapping of p m42,a new recessive wheat powdery mildew resistance gene derived fro m wild e mmer(Triticu m tur gidu m var.dicoccoides)[J].Theor Appl Genet,2009,119:223-230.
[15]Li G Q,Fang T L,Zhang H T,et al.Molecular identification of a new po wder y mildew resistance gene Pm41 on chr o moso me 3BL derived fro m wild e mmer(Triticu m t ur gidu m var.dicoccoides)[J].Theor Appl Genet,2009,119:531-539.
[16]Ghazaleh N,Volker M,Ger har d W,et al.Micr osatellite mapping of powdery mildew resistance allele Pm5d from common wheat line IGV1-455[J].Euphytica,2008,159:307-313.
[17]薛飛,翟雯雯,段霞瑜,等.小麥地方品種小白冬麥抗白粉病基因分子標記[J].作物學報,2009,35(10):1806-1811.
[18]Rong J K,Millet E,Manisterski J,et al.A new powder y mildew resistance gene:Introgression fro m wild emmer into common wheat and RFLP-based mapping[J].Euphytica,2000,115:121-126.
