棘阿米巴副衣原體病原學研究進展
2012-01-24 02:12:20吳移謀
中國人獸共患病學報 2012年11期
劉 彥,吳移謀
副衣原體科(Parachlamydiaceae)是1997年首次從2例女性阿米巴感染者鼻黏膜中分離出來的細胞內共生體,可觀察到原體、網狀體及新月體3種不同的發育期,大量證據表明它與人類呼吸道感染密切相關,還可能與動物的泌尿生殖道、眼部感染及全身性疾病有關,1999年被正式命名。副衣原體科主要包括棘阿米巴副衣原體(Parachlamydiaacanthamoeba,Pa)和哈氏變形蟲新衣原體(Neochlamydiaartmanellae,Nh)兩個屬。副衣原體屬目前僅有棘阿米巴副衣原體(P.acanthamoebae,Pa)一個單獨的種類,本文就棘阿米巴副衣原體分類、病原學特征及分離培養方法研究進展作一綜述。
1 棘阿米巴副衣原體分類
一個新的物種的鑒定,一般應該分析5種不同的種系進化。由于棘阿米巴副衣原體的基因序列尚未完全明確,目前主要是根據16SrRNA基因、23S rRNA基因、RNase P RNA基因及全基因的種系進化進行分類。Everett等通過對肺炎嗜衣原體(Chlamydophilapneumoniae)16SrRNA 基 因(GenBank登錄號為U68426)和/或23SrRNA基因(GenBank登錄號為U73784)的種系進化分析認為,與16SrRNA或23SrRNA基因同源性大于95% 的屬于衣原體科,反之則屬于副衣原體科。副衣原體科分為兩個屬,其典型菌株分別是棘阿米巴副衣原體和哈氏變形蟲新衣原體,目前已發現8個種的棘阿米巴副衣原體,分別為BN9、Bergl7、Hall’s球菌、UWE1、UWE25、UWC22、TUME1和 CorvenA4株。理論上認為,16SrRNA基因序列在同一屬的細菌中差別很小,同源性至少應該在97%以上;但經過比較,發現 UWE1、UWE25、UWC22、TUME1及CorvenA4等5株棘阿米巴副衣原體的16SrRNA基因與BN9株的同源性介于91% ~93%之間,說明棘阿米巴副衣原體在種系進化中具有呈多樣性。
2 棘阿米巴副衣原體病原學特征
2.1 形態與結構 棘阿米巴副衣原體可自然感染棘阿米巴的滋養體階段,是其細胞內共生體,復制周期與衣原體相似。電鏡下可觀察到原體(elementary body,EB)和網狀體(reticulate body,RB)兩種基本結構,有時可見新月體(crescent body,CB)(圖1)。原體和網狀體均可存在于棘阿米巴滋養體的空泡內,但革蘭染色結果不一致。
原體呈球形或橢球形,直徑為0.3~0.5μm,主要定位于棘阿米巴滋養體的空泡內,隨著培養時間的增加而增多,是副衣原體成熟階段,革蘭氏染色呈陽性。電鏡下可見原體細胞壁厚,有致密的類核結構和少量的核糖體(圖1A)。原體EB在外界環境較穩定,無繁殖能力,但具有高度的感染性;當進入棘阿米巴滋養體后,在其棘偽足包繞形成的空泡即吞噬空泡中逐漸發育,體積增大進入增殖狀態,形成始體。
網狀體呈球形,直徑稍大于原體,約為0.5~0.7μm,不同的菌株原體和始體直徑相差比較大,Hall’s coccus株始體直徑約0.6μm,BN9株直徑可達到1.0μm,CorvenA4株直徑為0.9μm。始體是副衣原體在棘阿米巴細胞質內的一個主要階段,但是在棘阿米巴的吞噬空泡中也數量眾多。始體呈二分裂相增殖,細胞壁薄,革蘭染色呈陰性。電鏡下可見始體電子密度低,無致密核質,內有纖細的網狀結構,又叫始體(initial body,IB)(圖1B)。
新月體與原體和網狀體共存,是原體和網狀體發育的中間階段,具有感染性,大量存在于外界環境中,還可寄生于棘阿米巴的吞噬空泡中,但在棘阿米巴胞漿內沒有發現;其主要功能可能是延長潛伏期。電鏡下可見新月體呈新月狀,大小約為0.3×0.6μm,細胞壁厚(圖1C),革蘭氏染色呈陽性。新月體發生機制尚不清楚,與共培養時間有關,隨著培養時間的延長,新月體出現頻率增加。新月體已見于副衣原體Berg17株、BN9和Hall’s coccus株,而衣原體均未見報道,因此新月體有可能用來作為區別衣原體分類的一個形態學特征。
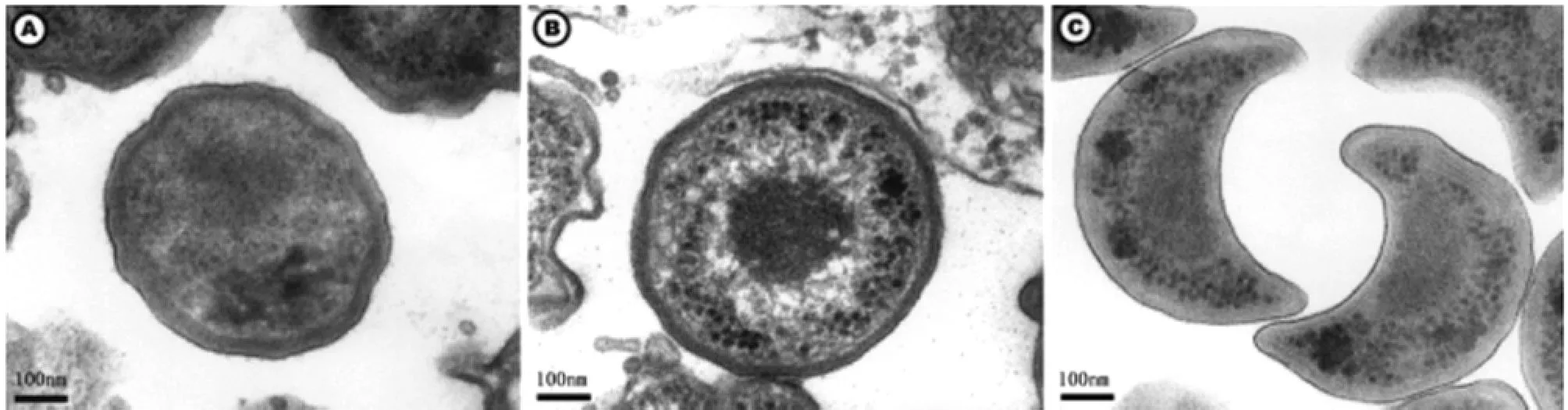
圖1 棘阿米巴副衣原體(Hall’s coccus株)3個不同階段的形態結構(電鏡,×45 600)A:原體;B:網狀體;C:新月體(Gilbert Greub and Didier Raoult,2002)Fig.1 Morphology of the three different stages of Parachlamydia acanthamoeba (Hall's coccus strain)(Electron microscopy,×45 600)(Greub G and Raoult D,2002)A:Elementary body;B:Reticulate body;C:Crescent body
2.2 發育周期 原體和新月體在外界環境性質均較穩定,富有感染性,可通過表面受體吸附于棘阿米巴滋養體,棘阿米巴伸出棘狀偽足將其吞入,并由自身胞膜包繞原體和新月體形成吞噬空泡(圖2);原體和新月體在吞噬空泡中逐漸發育、增大成為網狀體。有時可見散在的原體游離分布棘阿米巴胞漿中,而未見吞噬空泡,其形成來源可能為:①由體外經滋養體的胞吞作用進人胞漿中的原體,尚未開始形成吞噬空泡;②在成熟吞噬空泡中,發育成熟并具有感染能力的原體顆粒可發生胞內胞吐,釋放原體進入胞漿中,成為游離的原體。
網狀體在吞噬空泡內進行二分裂增殖,增殖后的網狀體可侵入棘阿米巴胞漿內,但不能繼續增殖,而吞噬空泡內的網狀體可繼續指數增殖,產生大量子代,形成致密的包涵體。網狀體在包涵體內逐漸發育成為原體或新月體,棘阿米巴滋養體的吞噬空泡體積增大,最終通過副衣原體的溶胞作用或者直接以胞吐方式釋放出來。棘阿米巴副衣原體的溶胞作用受溫度影響比較明顯,在低于30℃時表現較低的水平,而在32℃~37℃時達到最大值。
感染的棘阿米巴滋養體體積增加,出現空泡樣變化,可能是由于棘阿米巴副衣原體空泡的形成/或擴張;在培養后期,滋養體裂解,數量減少,大部分失去成囊活力,少部分輕度感染滋養體可形成包囊。據報道,Hall’s coccus株8h即可在多嗜棘阿米巴滋養體的吞噬空泡內發育成網狀體;滋養體發生空泡樣變化,可見分裂現象,胞漿內可見副衣原體包涵體,主要靠近細胞膜,呈瘤樣或球狀;滋養體體積增大,后期可見大部分滋養體裂解,數量減少;滋養體成囊率從99.6%下降至3%,囊內未發現存在副衣原體顆粒。在BN9株的研究中也發現,感染的多噬棘阿米巴滋養體的成囊能力下降,但在部分輕度感染的滋養體成囊后包囊內可發現副衣原體顆粒。
2.3 抗原結構 與衣原體相比,目前認為副衣原體的細胞壁中可能不具備脂多糖或N-末端截斷型脂多糖(truncated LPS);但副衣原體細胞壁上也含有一層蛋白質外膜,主要由幾種多肽組成,稱為主要外膜蛋白(major outer membrane protein,MOMP),它是決定其抗原性的主要物質基礎。
在衣原體的研究中發現,MOMP的二級結構與穿孔蛋白(porin)一致,具有穿孔蛋白特性。外膜蛋白A(outer membrane proteinA,OmcA)、外膜蛋白B(outer membrane protein B,OmcB)同 MOMP一起構成外膜復合物(outer membrane complex,OMC)。這些蛋白質的多態性非常明顯,從而決定了衣原體的多樣性。
經二維電泳及質譜分析,棘阿米巴副衣原體UWE25株已經發現38個外膜蛋白,蛋白pc0978、pc0616和pc0617富含半胱氨酸,可能與副衣原體細胞外膜的穩定性相關;其中pc0616和pc0617分別與衣原體OmcB、OmcA具備較高的同源性。pc1489和pc1077蛋白在副衣原體外膜中含量極高,研究已經證明,pc1489作為特異性抗體,定位于棘阿米巴副衣原體外膜[23],預測pc1489和pc1077的二級結構具備β-桶狀(β-barrel)結構,類似穿孔蛋白,被認為是副衣原體的MOMP;這兩種蛋白質與pc0978、pc0616和pc0617一起,構成了副衣原體的外膜復合物(圖3)。
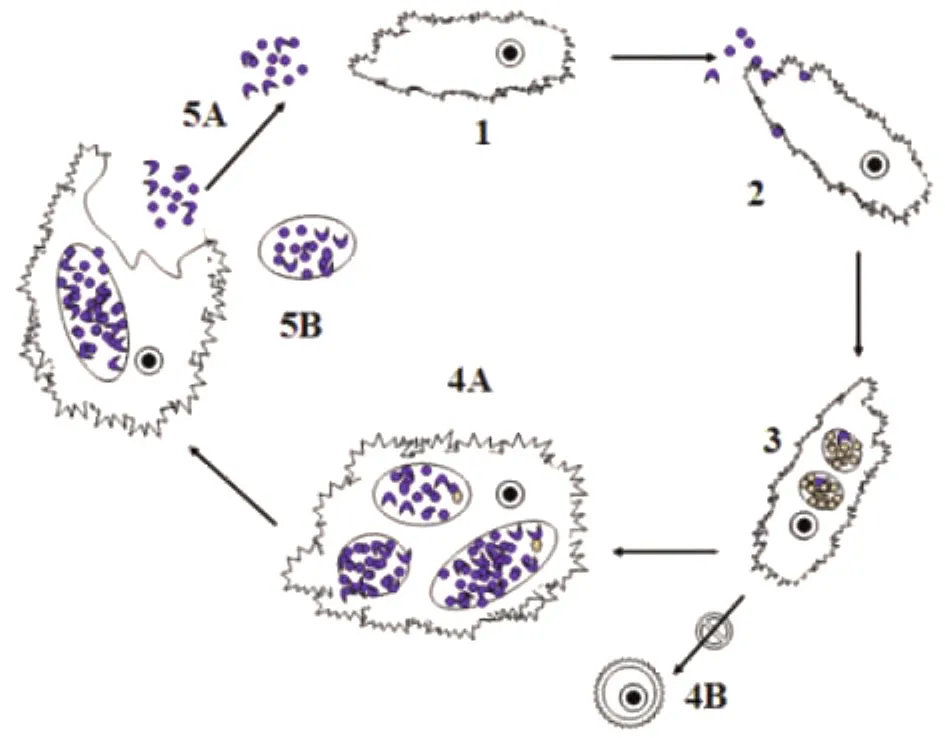
圖2 棘阿米巴副衣原體發育周期1、副衣原體(原體及新月體階段,紫色顆粒)與棘阿米巴(滋養體階段);2、棘阿米巴滋養體吞噬棘副衣原體;3、原體及新月體在棘阿米巴的吞噬空泡內體積增大形成網狀體(黃色顆粒),進行二分裂增殖,形成致密的包涵體;4、網狀體在包涵體內逐漸成熟為原體及新月體,棘阿米巴滋養體腫脹(4A),難以成囊(4B);5、棘阿米巴副衣原體以原體、新月體(5A)及小囊泡(5B)的形式釋放出來Fig.2 Developmental cycle of Parachlamydia acanthamoeba1:Parachlamydia (the elementary body and the crescent body,the purple particles)and Acanthamoeba(trophozoite);2:Acanthamoebatrophozoites swallowed the Parachlamydia;3:The elementary and crescent body turned to reticulate body(the yellow particles);4:The reticulate body turned to the elementary and crescent body,and the acanthamoebatrophozoites were difficult to turn to cysts;5:The Acanthamoebatrophozoites were split and the elementary and crescent body were released
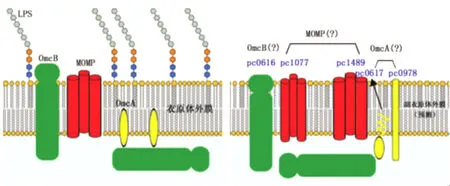
圖3 衣原體、副衣原體外膜基本結構模式圖左圖為衣原體外膜基本結構模式圖,右圖為副衣原體外膜基本結構預測圖。Fig.3 The diagram about structure of outer membrane of Chlamydiaand ParachlamydiaLeft:The structure of outer membrane of Chlamydia;Right:The predicted structure of Parachlamydia
目前已知的副衣原體UWE25株細胞壁外膜上的MOMP與已知衣原體沒有明顯同源性;這也進一步說明副衣原體UWE25和衣原體在細胞壁成分之間存在明顯的差異。
2.4 基因組結構與功能 目前只完成了副衣原體UWE25株全基因組測序(NC_005861)。UWE25基因組全長2 414 465bp,是衣原體基因數的2倍;其中存在3kb的gap,G+C含量為34.72%,比衣原體低得多;含有2 031個編碼區(Coding Sequence,CDS),938個CDS與其他衣原體具有同源性,其中711個CDS為副衣原體與其他衣原體所共有,而1 093個CDS只存在于UWE25中;副衣原體擁有3個rRNA操縱子,其數量多于其他衣原體的rRNA操縱子。
棘阿米巴副衣原體含有豐富的亮氨酸重復序列,采集了大量的植物類基因,并含有一個變形桿菌起源的大型基因組島Pam100G,包含有6個大型G+C 富 集 基 因 (large GC-rich genes,IgrA,IgrB,IgrC,IgrD,IgrE,IgrF),它們很可能起源于一個獨特的重復基因序列,與其他細菌未見任何同源性,在UWE25的生物學活性中發揮著重要的作用。由此可見,副衣原體的基因組遠遠大于其他衣原體的基因組,主要原因是其他衣原體生活在相對恒定的環境中(如真核細胞內),它們在進化過程中丟失了很多非必需的基因。
在 UWE25株基因島(genomic islands,GIS)研究中發現另一個G+C含量豐富的19kb的區域。這段序列是一個100kb的染色體的一部分,它包含100個高度保守的開放閱讀框(open reading frame,ORF),兩側有兩個17bp的重復序列,在這段遺傳密碼近段有兩個相同的GLY-tRNA串聯在一起;位于該染色體區域內有數個水平轉移獲得的基因,編碼轉座酶(transposases)及噬菌體相關蛋白,基本滿足GIS的標準。G+C含量分析表明,若干個序列組成這個基因組島(genomic island,GI),其中一組基因編碼了具有F-樣結構的DNA結合轉移系統,對F質粒的 DNA 接合傳遞 (traF,traG,traH,traN,traU,traW 及trbC)至關重要,說明副衣原體tra單位同樣參與了DNA的接合傳遞。這種F質粒的接合傳遞最有可能發生在感染了副衣原體的棘阿米巴內,涉及到副衣原體之間的DNA傳遞;這可能是副衣原體與衣原體遺傳學上的一個鑒別點;這些異源DNA很有可能是副衣原體從棘阿米巴的其他不同種的寄生物中獲取的,該基因島可能還是副衣原體一個重要致病物質。
同衣原體一樣,UWE25的合成能力有限,不能合成核苷酸和電子載體NAD+,只能通過一些特殊的渠道依靠宿主獲取,但能合成一些有限的氨基酸(甘氨酸,丙氨酸,絲氨酸,天門冬酸,谷氨酰胺,谷氨酸和脯氨酸)和輔助因子(血紅素,核黃素,葉酸和甲基萘醌類)等;這些特點證明了副衣原體專營寄生生活。
3 棘阿米巴副衣原體的分離培養
3.1 棘阿米巴中的副衣原體分離培養 副衣原體比較合適的自然宿主是棘阿米巴,極易與棘阿米巴共生,將其作為復制的載體,在棘阿米巴中復制增殖。迄今為止發現的8種副衣原體都寄生在人體或環境中的棘阿米巴中,因此副衣原體的培養主要依賴于棘阿米巴。目前可以從環境樣本和臨床標本檢測和分離獲得副衣原體,分離的樣品包括鼻拭子、支氣管肺泡灌洗液、感染的角膜組織、腦組織和皮膚病灶、粥樣硬化斑塊和外周血單個核細胞等,以及污水、土壤等;分離棘阿米巴中的副衣原體主要有兩種方法。
3.1.1 PYG分離培養:將環境樣本或臨床標本直接在蛋白胨酵母膏葡糖培養基(peptone yeast-extract glucose,PYG)中培養,PYG含有胰蛋白胨、酵母提取物及葡萄糖,營養豐富,必須添加雙抗(β-內酰胺類和喹諾酮類抗生素)和兩性霉素來去除污染。最佳生長溫度為28℃,需培養數天或幾周,培養過程中需要定期檢查有無棘阿米巴分解或Gimenez陽性球菌的出現[8]。
3.1.2 PAS分離培養:將環境樣本或臨床標本直接接種在PAS(Page’s modified Neff’s amoeba saline)培養基中培養,再覆蓋一層60℃熱滅活1hr的大腸桿菌(棘阿米巴以滅活的大腸桿菌為食)。PAS是一種專門用于阿米巴培養的培養基,其特點是培養基中無營養成分,能抑制臨床標本中污染物的過度生長,無須添加抗生素;于25~30℃培養,此法室溫下可保種3個月,4℃可保種6~12個月。A-strid等對工業廢水治理工廠中的活性淤泥(activated sludge)與未感染副衣原體的棘阿米巴標準株UWE1進行共同培養,成功獲得一株新的副衣原體命名為UV-7。經鑒定UV-7的16SrRNA序列與UWE1的同源性為98.7%。
3.1.3 研究證實抗生素、溫度和棘阿米巴狀態對副衣原體的分離培養有影響。
(1)抗生素:副衣原體對β-內酰胺類抗生素和喹諾酮類是耐藥的,而對慶大霉素較為敏感,使用此氨基甙類抗生素可阻礙副衣原體感染棘阿米巴;大環內酯類、四環素類抗生素、利福平及磺胺類藥物等也會抑制副衣原體的生長,故不能在PYG培養基中添加上述抗生素。
(2)溫度:棘阿米巴對生長條件的耐受性遠大于其它營自生生活阿米巴(如納氏蟲屬阿米巴),對干燥有較高抗性,對溫度要求也不嚴格,可在20~37℃ 左右很好的存活,但是在培養含有副衣原體的棘阿米巴時,孵育期間的溫度一般低于32℃,28℃是其最佳培養溫度。高溫下棘阿米巴非常容易形成包囊,而且在大于32℃的時候副衣原體易誘導阿米巴發生溶胞。冷凍可造成副衣原體的活力喪失,用液氮超低溫保存的方法,細胞的復蘇率極低,但添加2%~10% 胎牛血清可以減少失活率。
(3)宿主:副衣原體對外界的抵抗力主要源自于其寄生的棘阿米巴。一方面,它們的寄生宿主對溫度、pH值、滲透壓、消毒液以及抗生素等具有很強的耐受力,因此副衣原體對外界的抵抗力相對較強;另一方面,副衣原體與阿米巴的共生使它們互惠互利,副衣原體經過在阿米巴內的繁殖增強了傳播能力、耐藥性和致病性,同時也增強了自生生活阿米巴的抵抗力和致病性。
3.2 哺乳類細胞中的副衣原體分離培養 早期的研究認為副衣原體的宿主范圍受棘阿米巴屬限制,一般不用哺乳動物細胞培養副衣原體,但近來越來越多的實驗證明副衣原體菌株能在多種細胞中存活;1994年Michel等首次利用等猴Vero細胞成功培養了副衣原體Berg17株,從而開拓了在實驗室使用細胞培養副衣原體的新領域;Greub等發現副衣原體可以進入人體巨噬細胞,通過細胞內的大量復制,最后導致巨噬細胞凋亡[36]。體外細胞實驗證實,副衣原體感染肺泡細胞(A549)和肺成纖維細胞(HEL),并在細胞內進行復制;副衣原體BN9株在HEp-2和Vero細胞中也能很好地存活;副衣原體UV-7株不受宿主棘阿米巴UWE1的限制,可入侵哺乳動物細胞,如Vero,Hela和NCI-H292細胞。同其他衣原體一樣,棘阿米巴副衣原體利用ATP/ADP易位酶進行能量的轉換。棘阿米巴副衣原體通過細胞內吞的途徑進入人體巨噬細胞,并在其形成的吞噬空泡內進行大量復制,最后導致巨噬細胞的凋亡;副衣原體在巨噬細胞內的特征類似貝氏考克斯體(Coxiella burnetii)在內溶酶體內的特點,如呈酸性、溶酶體相關膜蛋白-1(Lamp-1)陽性及組織蛋白酶陰性;這些特點在本質上均不同于衣原體。
操作步驟包括棘阿米巴副衣原體樣本分離、感染宿主細胞及共同培養3部分。
(1)梯度離心-反復凍融法從棘阿米巴中分離副衣原體網狀體:感染副衣原體的棘阿米巴在PYG中32℃培養6d后,180×g離心10min去除棘阿米巴,取上清6 600×g離心30min后,PBS懸浮沉淀,加入含有10%蔗糖的PBS,4℃,5 800×g離心30min;不同濃度的泛影葡胺梯度超速離心后(140 000×g)收集棘阿米巴副衣原體,懸浮于PBS,在-80℃反復凍融兩次備用。
(2)感染宿主細胞:副衣原體網狀體經過DAPI(4,6-二脒基-2-苯基吲哚二鹽酸鹽,4′,6-diamidino-2-phenyl-indole,DAPI,DNA 熒光染料,可用于活細胞染色)染色后通過孔徑0.25μm的硝酸纖維素膜過濾并計數,或用倍比稀釋法計數。按1:1的比例,將宿主細胞與純化的棘阿米巴副衣原體(可100℃滅活1h)在RPMI培養基中37℃培養8h后,用RPMI-肝素鈉洗滌,繼續培養,無須添加抗生素。
(3)共同培養:感染的細胞在35℃,5%CO2條件下,培養3~5d,每天用PBS洗滌后,取部分細胞用1%甲醛固定10min,Diff-Quik法染色后觀察;也可培養3d后再進行DAPI染色觀察。
4 展 望
作為一個新發現的病原體,棘阿米巴副衣原體的致病作用需要得到進一步證實;因副衣原體的遺傳呈多樣性,不同的菌種其致病性也各不相同,因此它們的種系發生關系也需要進一步闡明;由于副衣原體難以培養,目前迫切需要從臨床標本和環境樣本中成功分離出棘阿米巴副衣原體菌株和/或發現未知的且同源性較高的新種類。分離培養出棘阿米巴副衣原體菌株,并對其進行分子生物學及血清學等相關研究,對進一步闡明其種系發生關系、證實其致病性、明確其致病機制并進行快速診斷均具有十分重大的意義。
[1]Amann R,Springer N,Schonhuber W,et al.Obligate intracellular bacterial parasites of acanthamoebae related to Chlamydia spp[J].Appl Environ Microbio,1997,63(1):115-121.
[2]Greub G,Boyadjiev I,La Scola B,et al.Serological hint suggesting that Parachlamydiaceae are agents of pneumonia in polytraumatized intensive care patients[J].Ann N Y Acad Sci,2003,990:311-319.
[3]Greub G,Berger P,Papazian L,et al.Parachlamydiaceae as rare agents of pneumonia[J].Emerg Infect Dis,2003,9(6):755-756.
[4]Greub G,Raoult D.Parachlamydiaceae:Potential emerging pathogens emerging[J].Infect Dis,2002,8(6):625-630.
[5]Collingro A,Toenshoff ER,Taylor MW,et al.Candidatus Protochlamydia amoebophila',an endosymbiont of Acanthamoeba spp[J].Int J Syst Evol Microbiol,2005,55(Pt 5):1863-1866.
[6]Fritsche TR,Horn M,Wagner M,et al.Phylogenetic diversity among geographically dispersed Chlamydiales endosymbionts recovered from clinical and environmental isolates of Acanthamoeba spp[J].Appl Environ Microbio,2000,66:2613-2619.
[7]Casson N,Posfay-Barbe KM,Gervaix A,et al.New diagnostic real-time PCR for specific detection of Parachlamydia acanthamoebae DNA in clinical samples[J].J Clin Microbiol,2008,46(4):1491-1493.DOI:10.1128/JCM.02302-07
[8]Thomas V,Casson N,Greub G.Criblamydia sequanensis,a new intracellular Chlamydialesisolated from Seine river water using amoebal co-culture[J].Environ Microbiol,2006,8:2125-2135.
[9]Greub G,Raoult D.Crescentbodies of Parachlamydia acanthamoeba and its life cycle within Acanthamoeba polyphaga:an electron micrograph study[J].Appl Environ Microbio,2002,68(6):3076-3084.
[10]Casson N,Michel R,Muller K,et al.Protochlamydia naegleriophila as etiologic agent of pneumonia[J].Emerg Infect Dis,2008,14(1):168-172.
[11]Greub G.Parachlamydia acanthamoebae,an emerging agent of pneumonia[J].Cell Mol Immunol,2009,15(1):18-28.DOI:10.1111/j.1469-0691.2008.02633.x
[12]Moelleken K,Hegemann JH.The Chlamydiaouter membrane protein OmcB is required for adhesion and exhibits biovar-specific differences in glycosaminoglycan binding[J].Mol Microbi-ol,2008,67(2):403-419.DOI:10.1111/j.1365-2958.2007.06050.x
[13]Eugster M,Roten CA,Greub G.Analyses of six homologous proteins of Protochlamydia amoebophila UWE25encoded by large GC-rich genes(lgr):a model of evolution and concatenation of leucine-rich repeats[J].BMC Evol Biol,2007,7:231.
[14]Greub G,Collyn F,Guy L,et al.A genomic island present along the bacterial chromosome of the Parachlamydiaceae UWE25,an obligate amoebal endosymbiont,encodes a potentially functional F-like conjugative DNA transfer system[J].BMC Microbiol,2004,48(4):1-12
[15]Boyer M,Yutin N,Pagniera I,et al.Giant marseillevirus highlights the role of amoebae as a melting pot in emergence of chimeric microorganisms[J].Proc Natl Acad Sci U S A,2009,106(51):21848-21853.DOI:10.1073/pnas.0911354106
[16]Greub G,Desnues B,Raoult D,et al.Lack of microbicidal response in human macrophages infected with Parachlamydia acanthamoebae[J].Microbes Infect,2005,7(4):714-719.
[17]Maurin M,Bryskier A,Raoult D.Antibiotic susceptibilities of Parachlamydia acanthamoebae in amoebae[J].Antimicrob A-gents Chemother,2002,46:3065-3067.
[18]Casson N,MedicoN,Bille J,et al.Parachlamydia acanthamoebae enters and multiplies with in pneumocytes and lung fibroblasts[J].Microbes Infect,2006,8(5):1294-1300.
[19]Matsuo J, Hayashi Y, Nakamura S,et al. Novel Parachlamydia acanthamoebae quantification method based on co-culturewith amoebae[J].Appl Environ Microbiol,2008,74(20):6397-6404.DOI:10.1128/AEM.00841-08
[20]Corsaro D,Greub G.Pathogenic potential of novel Chlamydiae and diagnostic approaches to infections due to these obligate intracellular bacteria[J].Clin Microbiol Rev,2006,19(2):283-297.
