紫扇貝和海灣扇貝雜交家系的生長和通徑分析
2012-01-11 14:42:06李建立王春德李朝霞蔡德泉張金盛
海洋科學(xué) 2012年8期
李建立, 王春德, 李朝霞, 蔡德泉, 張金盛
(1. 青島農(nóng)業(yè)大學(xué), 山東青島266109; 2. 威海市海洋與漁業(yè)局, 山東威海264200; 3. 招遠(yuǎn)市夏甸農(nóng)業(yè)技術(shù)推廣站, 山東煙臺 265415)
紫扇貝和海灣扇貝雜交家系的生長和通徑分析
李建立1, 王春德1, 李朝霞1, 蔡德泉2, 張金盛3
(1. 青島農(nóng)業(yè)大學(xué), 山東青島266109; 2. 威海市海洋與漁業(yè)局, 山東威海264200; 3. 招遠(yuǎn)市夏甸農(nóng)業(yè)技術(shù)推廣站, 山東煙臺 265415)
以紫扇貝(Argopecten purpuratus)和海灣扇貝(Argopecten irradians irradians)為親本, 建立了紫扇貝自交(ZZ)、海灣扇貝自交(HH)、紫扇貝(卵)×海灣扇貝(精)(ZH)和海灣扇貝(卵)×紫扇貝(精)(HZ)共 4個試驗家系。在養(yǎng)殖 60、127 d時測定殼高、殼長、殼厚和體質(zhì)量, 并分析各養(yǎng)殖時期影響體質(zhì)量的主要因素。結(jié)果表明: 在各養(yǎng)殖時期, 2雜交家系子代的殼高、殼厚以及體質(zhì)量均表現(xiàn)出生長優(yōu)勢(P<0.01), 因此認(rèn)為通過紫扇貝和海灣扇貝雜交方式是改善扇貝的良好方法。通徑分析的結(jié)果表明, 對于自交家系, 殼長是影響體質(zhì)量的主要影響因素, 但對于雜交家系HZ來說, 殼長(60 d)和殼厚(127 d)對體質(zhì)量的決定作用最大; 在雜交家系ZH, 60 d時殼長和殼厚的協(xié)同作用對體質(zhì)量的決定作用最大, 127 d時殼高的決定作用最大。因此對于兩個雜交家系的選育, 應(yīng)按照不同性狀對體質(zhì)量的決定系數(shù)選擇各時期的優(yōu)勢性狀, 以使子代的雜種優(yōu)勢充分表現(xiàn)。
紫扇貝(Argopecten purpuratus); 海灣扇貝(Argopecten irradians irradians); 雜交家系; 雜交優(yōu)勢; 通徑分析
海灣扇貝(Argopecten irradians irradians)自1982年引入中國以來深受養(yǎng)殖業(yè)戶的歡迎, 并形成了中國水產(chǎn)養(yǎng)殖業(yè)一個新的發(fā)展高潮[1]。但是, 近年來隨著養(yǎng)殖領(lǐng)域的逐漸擴大和養(yǎng)殖歷史的增長, 病害頻發(fā)、個體小型化等問題凸現(xiàn), 導(dǎo)致商品貝的規(guī)格越來越小。另外, 種質(zhì)退化也導(dǎo)致育苗養(yǎng)殖過程中較高的死亡率[2-3]。盡管從引種復(fù)壯等方面做了大量的工作,同時取得了一定的成效[4-8]。但是仍需要從種質(zhì)入手,借助于遺傳改良及育種技術(shù), 培養(yǎng)出一種抗逆性強、生長快、適應(yīng)性強的扇貝新品種。
紫扇貝(Argopecten purpuratus) 是原產(chǎn)于南太平洋的一種速生型中型扇貝[9]。紫扇貝的殼較寬, 閉殼肌肥大, 出肉率高, 且味道鮮美, 殼形優(yōu)美, 因此紫扇貝商品價值高。紫扇貝于2008年成功地從秘魯引進, 并成功地在家系水平進行了紫扇貝與海灣扇貝的種間雜交實驗[10-11]。體質(zhì)量是扇貝的主要經(jīng)濟性狀之一, 是選擇育種的一個重要指標(biāo), 同時也是所有經(jīng)濟性狀的綜合體現(xiàn)。因此, 找出影響體質(zhì)量的主要目標(biāo)性狀, 明確選育目的, 具有重要的現(xiàn)實意義。近年來, 多元回歸和通徑分析已在水產(chǎn)動物魚類、蝦類和貝類研究中得到廣泛應(yīng)用, 研究內(nèi)容主要集中在生長性狀對體質(zhì)量的影響[12-18]。為了獲得通過遺傳改良的新品系, 2009年, 采用同樣的方法建立了1個紫扇貝(卵)×海灣扇貝(精)雜交家系和1個海灣扇貝(卵)×紫扇貝(精)雜交家系。通過比較2個雜交家系與紫扇貝和海灣扇貝自交家系之間經(jīng)濟性狀的差異, 并通過通徑分析方法, 確定家系中各形態(tài)性狀對體質(zhì)量的相對重要性, 以滿足生產(chǎn)的需要。
1 材料與方法
1.1 實驗材料
2009年 5月, 采用紫扇貝與海灣扇貝種間雜交的方法[10], 獲得兩個雜交家系和兩個自交家系。
ZH: 紫扇貝(卵)×海灣扇貝(精);
HZ: 海灣扇貝(卵)×紫扇貝(精);
ZZ: 紫扇貝(卵)×紫扇貝(精);
HH: 海灣扇貝(卵)×海灣扇貝(精)。
親本的規(guī)格見表 1。分別隔離培養(yǎng) 4個家系,幼蟲培育過程中控制相同密度; 幼蟲附著后, 及時分苗并控制各組密度; 2009年7月, 各組扇貝的養(yǎng)成過程均在青島小口子自然海區(qū)扇貝籠中進行,每層30只(每籠10層), 養(yǎng)成實驗至 2009年 11月底結(jié)束。

表1 實驗親本的大小和體質(zhì)量Tab.1 The morphological measurements of brood stocks (n=50)
1.2 實驗方法
在養(yǎng)成階段的第60天和第127天, 分別從各實驗家系中隨機抽取50只扇貝, 用游標(biāo)卡尺測量每只扇貝的殼高、殼長和殼厚, 電子天平秤量扇貝的體質(zhì)量。
1.3 分析方法
相關(guān)數(shù)據(jù)分析采用 SPSS 13.0 統(tǒng)計分析軟件處理。對于數(shù)據(jù)比較結(jié)果, 當(dāng)P<0.05 時為差異顯著,P<0.01時為差異極顯著。用下面的公式來計算雜種優(yōu)勢(Heterosis):
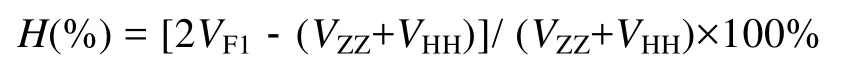
公式中的 VF1代表雜交子一代(ZH或 HZ)性狀的平均值,VZZ和VHH分別代表ZZ和HH性狀的平均值。計算各性狀平均數(shù)和標(biāo)準(zhǔn)差, 獲得各性狀表型參數(shù)統(tǒng)計量, 對各生長參數(shù)進行方差分析、顯著性檢驗和Duncan 多重比較分析、各性狀間表型相關(guān)分析和形態(tài)性狀各指標(biāo)對體質(zhì)量的通徑分析。在表型相關(guān)分析的基礎(chǔ)上, 根據(jù)通徑系數(shù)原理得到各形態(tài)性狀對體質(zhì)量的通徑系數(shù), 分析各性狀分別對體質(zhì)量的直接和間接影響, 并求得決定系數(shù)[12]。
2 結(jié)果
2.1 子代的生長性狀比較
各組在60 d 時生長性狀的生長情況見表2, 兩個雜交家系(HZ和ZH)之間的殼高、殼長、殼厚和體質(zhì)量差異均不顯著(P>0.05), 而ZH和HZ的殼高、殼長、殼厚和體質(zhì)量極顯著高于ZZ(P<0.01), HH的體質(zhì)量和ZH與HZ相比差異不顯著(P>0.05), HH的殼高、殼長、殼厚和體質(zhì)量極顯著高于ZZ(P<0.01)。各組在127 d時生長性狀的生長情況見表3, ZH的殼高、殼長、殼厚和體質(zhì)量和 HZ相比差異不顯著(P>0.05), ZH和HZ的殼高、殼長、殼厚和體質(zhì)量極顯著高于ZZ和HH(P<0.01), HH的殼高、殼長、殼厚和體質(zhì)量極顯著高于ZZ(P<0.01)。
2個雜交家系的雜種優(yōu)勢率見表3, 隨著養(yǎng)成時間的增加, ZH和HZ各生長性狀的雜種優(yōu)勢率有增大的趨勢。在60 d和127 d , ZH和HZ的雜種優(yōu)勢率大小依次是體質(zhì)量>殼長>殼高>殼寬。在 60 d時, ZH的體質(zhì)量雜種優(yōu)勢率最大, 是32.37%, HZ的體質(zhì)量雜種優(yōu)勢率是27.75%, ZH的體質(zhì)量和殼高的雜種優(yōu)勢率高于HZ。在127 d 時, ZH的體質(zhì)量雜種優(yōu)勢率最大, 是77.95%, 比HZ的高, ZH殼高、殼長和殼寬的雜種優(yōu)勢率低于HZ。
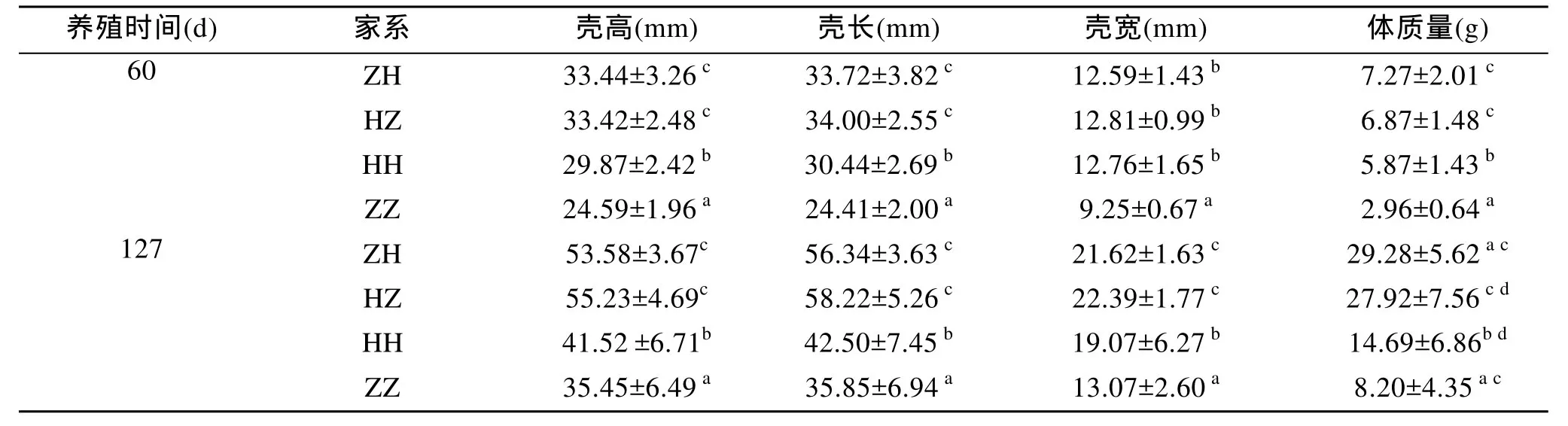
表2 各子代家系的生長指標(biāo)Tab. 2 The growth parameters of the hybrid and purebred families
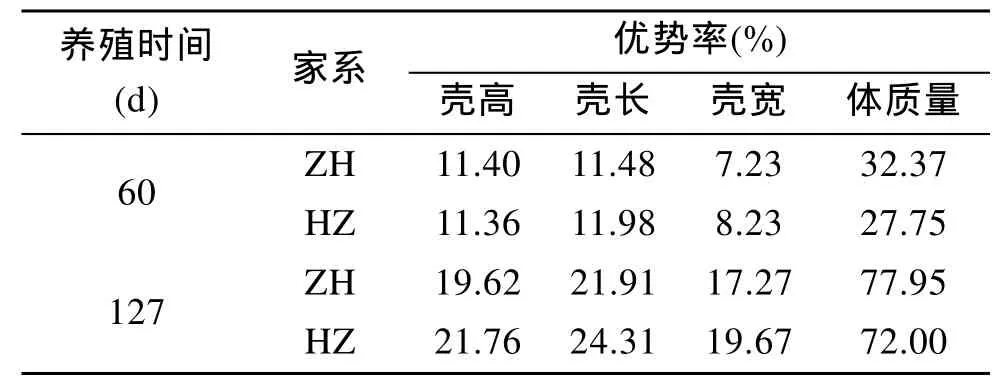
表3 雜交家系的雜種優(yōu)勢率Tab. 3 The heterosis of hybrid families
2.2 各組主要生長性狀對體質(zhì)量的通徑分析
通過相關(guān)分析, 3個生長性狀與體質(zhì)量的相關(guān)系數(shù)見表4。4個家系在60 d時的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼長>殼高>殼寬; 在127 d時, ZH和HH的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼長>殼高>殼寬, HZ的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼寬>殼高>殼長, ZZ的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼寬>殼長>殼高。
根據(jù)通徑分析原理, 得到各性狀對活體質(zhì)量的通徑系數(shù)(表 4), 根據(jù)相關(guān)系數(shù)的組成效應(yīng), 可將貝殼各性狀與活體質(zhì)量的相關(guān)系數(shù)剖分為各性狀的直接作用Pi和各性狀通過其他性狀的間接作用兩部分, 即rxiy=Pi+rijPj, 結(jié)果見表4。通徑系數(shù)反映自變量對依變量的直接影響, 結(jié)果表明 60 d時, 對于自交家系HH和ZZ殼長對體質(zhì)量的直接作用最大, 雜交家系ZH和HZ殼長對體質(zhì)量的直接作用最大。4組殼高殼長和殼寬對體質(zhì)量的間接作用大于直接作用, 主要是殼高和殼寬對體質(zhì)量產(chǎn)生的間接作用。在127 d時, 對于自交家系HH殼長對體質(zhì)量的直接作用最大, ZZ殼寬對體質(zhì)量的直接作用最大, HH和ZZ殼高、殼長和殼寬對體質(zhì)量的間接作用大于直接作用, 主要是殼高和殼寬對體質(zhì)量產(chǎn)生的間接作用。雜交家系ZH殼高對體質(zhì)量的直接作用最大, ZH的殼高、殼長和殼寬對體質(zhì)量的直接作用大于間接作用。雜交家系HZ殼寬對體質(zhì)量的直接作用最大, HZ殼高、殼長和殼寬對體質(zhì)量的間接作用大于直接作用,主要是殼高和殼長對體質(zhì)量產(chǎn)生的間接作用。
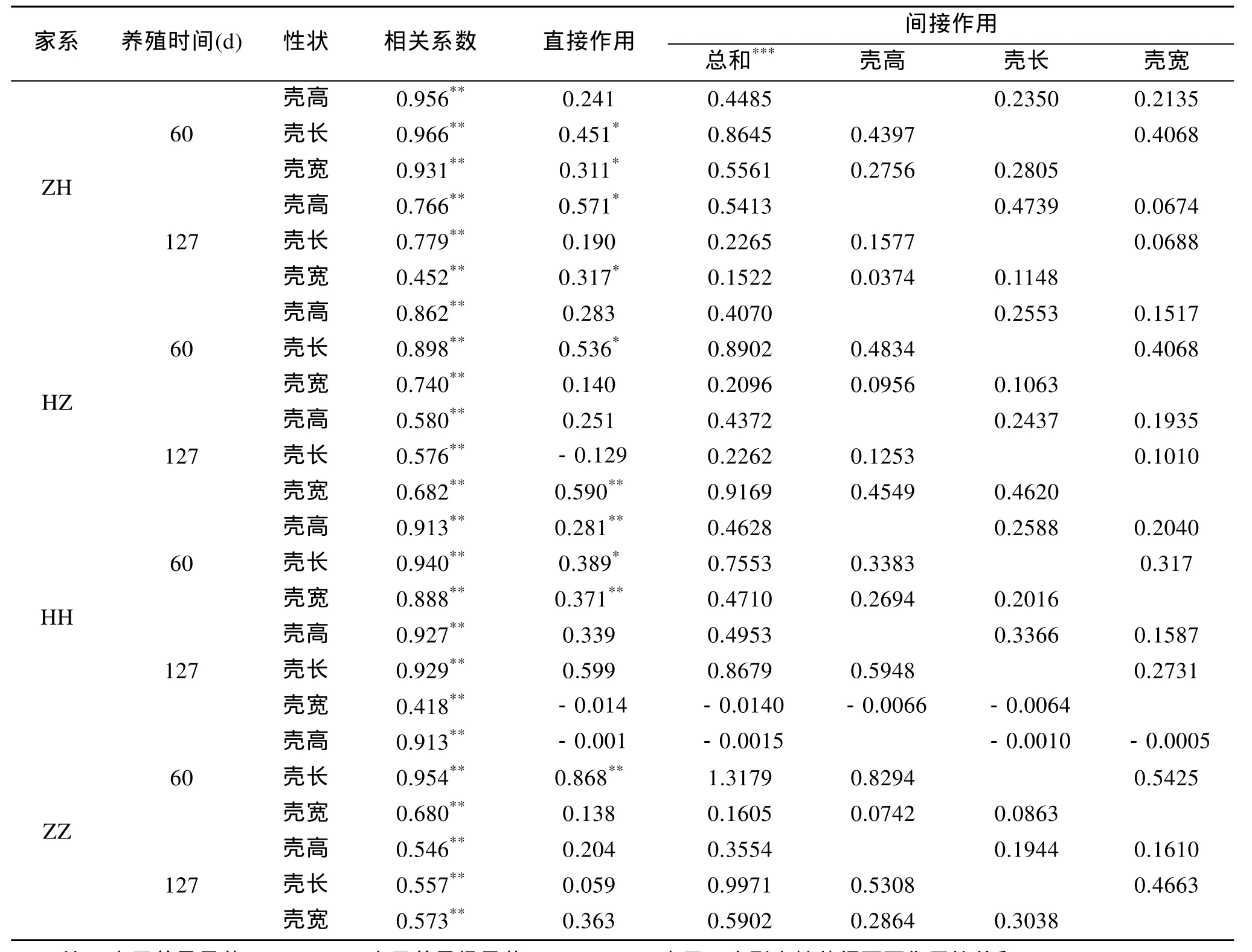
表4 各性狀對活體質(zhì)量的影響的通徑分析Tab. 4 Path analysis of the effects of the economic traits on body weight
2.3 各組主要生長性狀對體質(zhì)量的決定程度
根據(jù)單個性狀對活體質(zhì)量的決定系數(shù)為Di=Pi2,兩個性狀對活體質(zhì)量的共同決定系數(shù)Dij=2rijPiPj,計算出貝殼各性狀間協(xié)同對活體質(zhì)量的決定系數(shù)見表 5。對于雜交家系 ZH, 殼長和殼寬(60 d)的共同決定作用最大, 殼高(127 d)單獨決定作用最大;雜交家系 HZ, 殼長(60 d)的單獨決定作用最大, 殼寬(127 d)的單獨決定作用最大; 自交家系HH殼長和殼寬(60 d)的共同決定作用最大, 殼高和殼長(127 d)共同決定作用最大; 自交家系ZZ殼長(60 d)的單獨決定作用最大, 殼寬(127 d)的單獨決定作用最大。
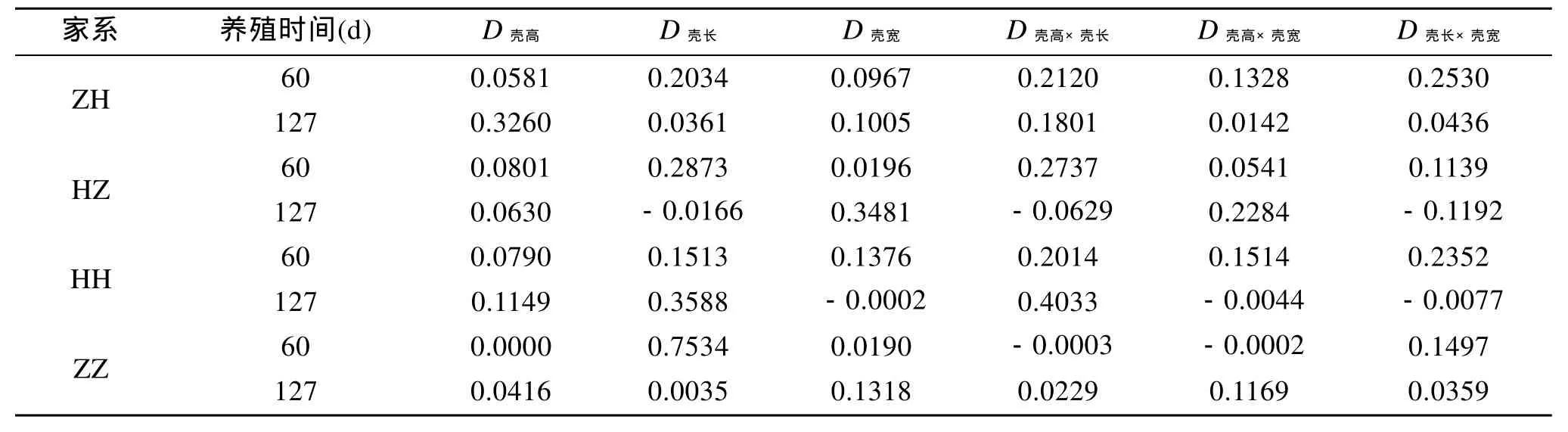
表5 貝殼各性狀對活體質(zhì)量的決定系數(shù)Tab. 5 The determinant coefficients of the traits on body weight
2.4 多元回歸方程的建立
根據(jù)多元相關(guān)和通徑分析的結(jié)果, 以活體質(zhì)量為依變量, 以對其產(chǎn)生顯著影響的其他主要性狀為自變量進行了多元回歸分析,X1,X2,X3分別是殼長、殼高和殼多元回歸方程寬, 建立以殼長、殼高、殼寬為自變量, 活體質(zhì)量為依變量的多元回歸方程, 各組的多元回歸方程見表6。結(jié)果顯示, 各組線性回歸關(guān)系達(dá)到極顯著水平(P<0.01)。

表6 多元回歸方程Tab. 6 Multiple regression equations
3 討論
3.1 各組生長比較
海灣扇貝引種養(yǎng)殖的成功極大地推動了中國海水養(yǎng)殖業(yè)的發(fā)展, 但是, 進入20 世紀(jì)90 年代以后,隨著海灣扇貝全人工育苗養(yǎng)殖代數(shù)的增加, 在生產(chǎn)中逐漸出現(xiàn)一些問題, 例如生長速度減緩、病害頻發(fā)和商品規(guī)格小型化等。產(chǎn)生這些問題的原因與海區(qū)環(huán)境惡化、養(yǎng)殖結(jié)構(gòu)不合理、病原生物增加等有關(guān),也與連續(xù)多代近交可能導(dǎo)致的近交衰退、抗逆能力減弱等引起種質(zhì)質(zhì)量下降有關(guān)。盡管通過養(yǎng)殖技術(shù)和反復(fù)引種都能夠使上述問題得到一定程度的解決,但是無法從根本上解決問題, 只有借助于遺傳育種技術(shù)提高其抗逆性和培育新品種, 才能使這些問題從根本上得到解決[19-21]。在養(yǎng)成實驗中發(fā)現(xiàn), 紫扇貝和海灣扇貝的雜交子代在殼高、殼長、殼寬和體質(zhì)量方面和海灣扇貝與紫扇貝相比均表現(xiàn)出較高的生長優(yōu)勢(P<0.01)。在60 d和127 d時, 2個雜交家系的4個經(jīng)濟性狀具有一定的雜種優(yōu)勢, 2個雜交家系的殼高、殼長、殼寬和體質(zhì)量的雜種優(yōu)勢率隨著養(yǎng)成時間的增加而逐漸增大。在60 d, 在4個經(jīng)濟性狀中, ZH和HZ體質(zhì)量雜種優(yōu)勢率最大, ZH的殼高和體質(zhì)量雜種優(yōu)勢率高于HZ, 殼長、殼高和殼寬也表現(xiàn)出一定的雜種優(yōu)勢率, 這表明紫扇貝和海灣扇貝雜交改良扇貝品質(zhì)是可行的。
3.2 通徑分析
方差分析的結(jié)果表明, 在各養(yǎng)殖時期, 四個家系的形態(tài)性狀和體質(zhì)量均存在顯著極差異(P<0.01),根據(jù)這一結(jié)果來確定影響體質(zhì)量的優(yōu)勢形態(tài)性狀是很困難的。在動物育種中, 利用不同性狀之間的相關(guān)性可為選育工作提供方便。但是簡單相關(guān)分析不能全面考察變量間的相互關(guān)系, 使結(jié)果帶有一定的片面性。多元回歸分析在一定程度上能夠消除變量之間的混淆, 但由于偏回歸系數(shù)帶有單位, 使原因?qū)Y(jié)果的效應(yīng)不能直接進行比較。生長性狀除通過直接作用引起目標(biāo)性狀變化外, 還可通過其他性狀以間接作用引起目標(biāo)性狀變化[22]。Wright于1921年提出了通徑分析方法, 該方法克服了相關(guān)分析與回歸分析的不足, 能夠真實反映各自變量和依變量的關(guān)系, 而且由于通徑系數(shù)是變量標(biāo)準(zhǔn)化的偏回歸系數(shù),從而使原因?qū)Y(jié)果的效應(yīng)能直接進行比較[22]。同時通徑系數(shù)還能區(qū)分原因?qū)Y(jié)果的直接作用和間接作用, 能全面反映原因和結(jié)果的相對重要性。由于通徑分析把相關(guān)系數(shù)剖分成兩部分, 其中直接影響部分即是通徑系數(shù), 且不受其他有關(guān)變量的影響, 因此可以通過通徑分析來探明兩個性狀之間真正關(guān)系[23]。
通過相關(guān)分析, 4個家系在60 d時的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼長>殼高>殼寬; 在127 d時, ZH和HH的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼長>殼高>殼寬, HZ的 3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼寬>殼高>殼長, ZZ的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼寬>殼高>殼長。在127 d時, ZH殼高和殼長的直接作用大于間接作用, 直接作用最大的是殼高, 在60 d時, 3個生長性狀的間接作用均大于直接作用; 其他家系在兩個生長時期內(nèi) 3個生長性狀的間接作用均大于直接作用。例如, 在127 d時,ZH的3個生長性狀與體質(zhì)量的相關(guān)系數(shù)大小依次為殼長>殼高>殼寬, 它的通徑系數(shù)大小依次為殼高>殼寬>殼長。因此, 采用通徑分析不僅可以分析計算出各個自變量與依變量之間的直接關(guān)系和間接關(guān)系,并且可以發(fā)現(xiàn)各個自變量之間存在相互制約關(guān)系,才產(chǎn)生了相關(guān)系數(shù)大而通徑系數(shù)不一定大的現(xiàn)象。劉小林等[15]在櫛孔扇貝的研究中也發(fā)現(xiàn)了這一現(xiàn)象。本實驗數(shù)據(jù)所建立的多元回歸方程, 經(jīng)檢驗, 回歸關(guān)系極顯著, 偏回歸關(guān)系也極顯著, 表明該方程具有較廣泛的適用范圍和實用價值。
根據(jù)本文中通徑分析和決定系數(shù)分析的結(jié)果來看, 以127 d為標(biāo)準(zhǔn), 在對高活體質(zhì)量雜交扇貝的選育過程中, ZH家系綜合起來看應(yīng)優(yōu)先考慮的性狀為殼高(直接作用大于間接作用), 其次是殼寬和殼長,殼高、殼寬和殼長均為易測性狀, 且殼高對活體質(zhì)量的通徑系數(shù)和總的決定系數(shù)在所有形態(tài)性狀中最大,是影響活體質(zhì)量的最重要的形態(tài)性狀, 因此, 其可以作為第一目標(biāo)性狀加以選育, 同時, 可把殼寬作為第二、把殼長作為第三目標(biāo)性狀對殼高加以輔助選育, 以達(dá)到最佳的選育效果。而HZ家系的3個性狀的間接作用大于直接作用, 主要是殼高和殼長對體質(zhì)量產(chǎn)生的間接作用, 但是比較 3個性狀的直接作用發(fā)現(xiàn), 殼寬對體質(zhì)量的直接作用最大, 可以作為第一目標(biāo)性狀, 同時, 殼高和殼寬做為第二和第三目標(biāo)性狀對殼寬加以輔助選育。
[1]張福綏, 何義朝, 元玲欣, 等. 墨西哥灣扇貝的引種和子一代苗種培育[J]. 海洋與湖沼, 1994, 25(4):272-377.
[2]張福綏. 近現(xiàn)代中國水產(chǎn)養(yǎng)殖業(yè)發(fā)展回顧與展望[J].世界科技研究與發(fā)展, 2003, 25(3): 5-13.
[3]李云福, 劉路偉, 邢光敏, 等. 美國海灣扇貝引種制種及選育技術(shù)報告[J]. 河北漁業(yè), 2000, 2: 29-32.
[4]張福綏. 海灣扇貝引種復(fù)壯研究[J]. 海洋與湖沼,1997, 28(2): 146-152.
[5]張國范, 劉述錫, 劉曉, 等. 海灣扇貝自交家系的建立和自交效應(yīng)[J]. 中國水產(chǎn)科學(xué), 2003, 10(6):441-445.
[6]鄭懷平, 張國范, 劉曉, 等. 不同貝殼顏色海灣扇貝(Argopecten irradians)家系的建立及生長發(fā)育研究[J].海洋與湖沼, 2003, 34(6): 632-638.
[7]鄭懷平, 張國范, 劉曉, 等. 海灣扇貝雜交家系與自交家系生長和存活的比較[J]. 水產(chǎn)學(xué)報, 2004, 28(3):267-272.
[8]秦艷杰, 劉曉, 張海濱, 等. 海灣扇貝雜交后代的微衛(wèi)星鑒定[J]. 中國水產(chǎn)科學(xué), 2007, 14(4): 672-677.
[9]Gonzalez M L, Perez M C, Lopez D A. Breeding cycle of the northern scallopArgopecten purpuratus(Lamarck, 1819) in southern Chile [J]. Aquaculture Res,2002, 33: 847-852.
[10]王春德, 劉保忠, 李繼強, 劉升平. 紫扇貝與海灣扇貝種間雜交的研究[J]. 海洋科學(xué), 2009, 33(10):83-91.
[11]Wang C, Liu B, Li J, et al. Introduction of the Peruvian scallop and its hybridization with the bay scallop in China[J]. Aquaculture, 2011, 310: 380-387.
[12]佟雪紅, 董在杰, 繆為民, 等. 建鯉與黃河鯉的雜交優(yōu)勢研究及主要生長性狀的通徑分析[J]. 大連水產(chǎn)學(xué)院學(xué)報, 2007, 22(3): 159-163.
[13]Turker H, Eversole A G. Evaluation of nondestructive method for determining body composition of crayfish[J].Journal of Shellfish Research, 1998, 17(1): 339.
[14]董世瑞, 孔杰, 萬初坤, 等. 中國對蝦形態(tài)性狀對體重影響的通徑分析[J]. 海洋水產(chǎn)研究, 2007, 28(3):15-22.
[15]劉小林, 常亞青, 相建海, 等. 櫛孔扇貝殼尺寸性狀對活體重的影響效果分析[J]. 海洋與湖沼, 2002,33(6): 673-678.
[16]劉志剛, 王輝, 孫小真等. 馬氏珠母貝經(jīng)濟性狀對體重決定效應(yīng)分析[J]. 廣東海洋大學(xué)學(xué)報, 2007, 27(4):15-20.
[17]何毛賢, 史兼華, 林岳光, 等. 馬氏珠母貝生長性狀的相關(guān)分析[J]. 海洋科學(xué), 2006, 30(11): 1-4.
[18]袁志發(fā), 周敬芋, 郭滿才. 決定系數(shù)—通徑系數(shù)的決策指標(biāo)[J]. 西北農(nóng)林科技大學(xué)學(xué)報(自然科學(xué)版),2001, 29(5): 131-133.
[19]張福綏, 何義朝, 亓玲欣, 等. 海灣扇貝引種復(fù)壯研究[J]. 海洋與湖沼, 1997, 28(2): 146-152.
[20]張國范, 李霞, 薛真福. 我國貝類大規(guī)模死亡的原因分析及防治對策[J]. 中國水產(chǎn), 1999, 9: 34-39.
[21]Chew K K. Global bivalve shellfish introductions [J].World Aquaculture, 1990, 21: 9-22.
[22]李朝霞, 王春德. 海灣扇貝自交與雜交子代的生長比較和通經(jīng)分析[J]. 中國農(nóng)學(xué)通報2009, 25(8): 282-285.
[23]盛志廉, 吳常信. 數(shù)量遺傳學(xué)[M]. 北京: 中國農(nóng)業(yè)出版社, 1999: 16-26.
Growth and path analyses of hybrid families between the Peruvian and bay scallop
LI Jian-li1, WANG Chun-de1, LI Zhao-xia1, CAI De-quan2, ZHANG Jin-sheng3
(1. Qingdao Agricultural University, Qingdao 266109, China; 2. Weihai Bureau of Ocean and Fisheries, Weihai 264200, China; 3. Xiadian Agricultural Technology Promotion Station, Zhaoyuan 265415, China)
Nov. ,18, 2011
Argopecten purpuratus;Argopecten irradians irradians; hybrid family; heterosis; path analysis
Two inter-specific hybrid families (HZ and ZH) and two inbred families (ZZ and HH) were constructed using the Peruvian scallop (Argopectenpurpuratus) and the bay scallop (Argopecten irradians irradians). Their shell height, shell length, shell width and whole weight were measured on Day 60 and 127 and the major factors affecting their body weight were analyzed. The results showed obvious growth hetereosis in shell height, shell thickness and whole weight (P<0.01). Path analyses showed that shell length was the major factor affecting the whole weight for inbred families. While for the hybrid family HZ, shell length on Day 60 and shell width on Day 127 were the major factors affecting whole weight. In the hybrid family ZH, the interaction between shell length and shell width on Day 60 and the shell height on Day 127 were the major factors affecting whole body weight.Therefore, different parameters should be used in selection of the hybrid families at different stages to ensure the greatest hetereosis.
S917.3
A
1000-3096(2012)08-0015-06
2011-11-18;
2012-04-26
國家自然科學(xué)基金項目(31172404); 國家自然科學(xué)青年基金(30901101); 農(nóng)業(yè)部引進國際先進農(nóng)業(yè)科學(xué)技術(shù)(948)項目(2008-Z31,2011-Z27); 青島市科技計劃項目(11-2-1-14-hy)
李建立(1987-), 男, 碩士研究生, 主要從事海洋生物遺傳育種學(xué)研究, E-mail: lijianli008@126.com; 王春德, 通信作者, 博士, 主要從事海洋生物遺傳育種學(xué)研究, E-mail: chundewang2007@163.com
梁德海)
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
小讀者(2021年2期)2021-03-29 05:03:48
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年2期)2019-08-23 08:12:08
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
