普通PCR與TD-PCR擴增葉爾羌高原鰍抗菌肽Hepcidin基因的比較
2011-12-10 10:35:02劉書東王娟娟陳根元李蓮瑞
塔里木大學學報 2011年4期
劉書東 王娟娟 陳根元 李蓮瑞*
(1 塔里木大學生命科學學院,新疆阿拉爾 843300)
(2 塔里木畜牧科技兵團重點實驗室,新疆阿拉爾 843300)
葉爾羌高原鰍[triplophysa(Hedinichthys)yarkandensis(Day)]為鰍科、高原鰍屬、鼓鰾亞屬的魚類,俗名葉爾羌條鰍、狗頭魚[1,2]。主要分布新疆南部的塔里木河水系,但近年來,由于環境污染、病害頻繁,造成了資源的缺乏,成為瀕危魚類。對葉爾羌高原鰍病害的防治,除了保護好當地的環境,還可以提高魚本身的免疫能力。在病害早期,魚類主要依靠非特異性體液免疫來抵御病原體感染,而在此過程中抗菌肽起到一定的重要作用。(Antibacterial peptides)是一類由基因編碼的小分子多肽,是機體免疫防衛系統的重要組成部分,生物體產生的對抗外源性病原體侵襲致病作用的防御性肽類活性物質,一般由10~50個氨基酸組成[3]。抗菌肽分子量小,活性強,功能廣泛,應用基因工程克隆與表達抗菌肽基因,改造合成抗菌肽基因以及動物的轉抗菌肽基因工程等已越來越受到人們的重視。Hepcidin作為魚類抗菌肽的一種類型備受關注,是一種有肝臟特異表達的抗菌肽,相對分子質量小(4 kD左右),具有顯著的廣譜抗菌活性,是機體天然免疫的一種效應子,也是調節鐵代謝的重要分子,目前被認為是調節維持鐵穩態極其重要的負激素[4]。降落PCR(touchdown PCR,TD—PCR)是一種設計多循環以使相連循環的退火溫度越來越低,從而達到最佳擴增條件的方法。本實驗利用降落式PCR技術擴增葉爾羌高原鰍抗菌肽基因特定片段,獲得良好效果。
1 材料和方法
1.1 材料
1.1.1 實驗動物:
葉爾羌高原鰍(Triplophysa(Hedinichthys)yarkandensis(Day))試驗用魚,系塔里木河流域阿拉爾河段,人工小抬網和地籠捕撈而得。
1.1.2 儀器與試劑:電泳儀DYY-12穩壓電泳儀、TC-512型PCR儀、凝膠成像儀;PCR擴增所用的主要試劑dNTPs、TaqDNA酶、DL 2000 Marker購自TakaRa、超純凈水;瓊脂糖由北京百泰克生物技術有限公司生產。
1.2 試驗方法
1.2.1 總RNA的提取:
本研究參照徐麗君等提取葉爾羌高原鰍肝胰臟總RNA的方法進行提取[5]。
1.2.2 瓊脂糖凝膠電泳
采用分子克隆[6]方法進行。
1.2.3 第一條鏈合成
取 RNA8 μL,dNTP 1 μL ,OligdT 1 μL 65 ℃,5 min,冰上1~2 min;在分別加入5×Primesoript buffer 4 μL,RNA Inhibiter 0.5 μL,Primer script RTase1 μL,PCR H2O,混勻,42 ℃ 45 min,70 ℃ 15 min,冰浴。
1.2.4 引物設計
根據 GenBank上已報道的魚類Hepcidin基因序列設計引物:
PF:5'-ggcGAATTCATGAAGACATCAGTGTTTGCTG-3'
PR:5'-ggcCTCGAGTAGTGGTCACAGGACCCGTCA-3'
引物由北京華大中生科技發展有限公司合成,引物PF和PR用無RNA酶水稀釋成20 pmol/μL的工作濃度備用。
1.2.5 普通PCR反應體系與反應程序:
普通PCR反應體系(20 μL)及反應程序:PCR反應體系:dNTP 2 μL,d3H2O 12.5 μL,10 × Buffer 2 μL, PF0.5 μL, PR0.5 μL, DNA 1 μL,Taq 0.4 μL。
反應程序為:94℃預變性5 min;94℃變性l min;59.5℃退火30 s;72℃延伸l min進行32個循環;72℃延伸10 min。
PCR擴增結束,取5 μL擴增產物于l%瓊脂糖凝膠,100 V電泳20 min,用VP凝膠成像系統觀察并成像分析。
1.2.6 降落PCR反應體系與反應程序:
PCR 反應體系:dNTP 2 μL,d3H2O 12.5 μL,10 × Buffer 2 μL,PF 0.5 μL,PR 0.5 μL,DNA 1 μL,Taq 0.4 μL;PCR 擴增程序:94℃ 5 min;94℃30 s,63 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,62℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,61 ℃ 30 s,72 ℃ 90 s,2次循環;94 ℃ 30 s,60 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,59 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,58 ℃ 30 s,72 ℃ 90 s,2次循環;94℃ 30 s,57 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s,2 次循環;94 ℃30 s,54 ℃ 30 s,72 ℃90 s,2 次循環;94 ℃ 30 s,53 ℃ 30 s,72 ℃ 90 s,2次循環;94 ℃ 30 s,52 ℃ 30 s,72 ℃ 90 s,2 次循環;94℃30 s,55 ℃ 30 s,72 ℃ 90 s,26次循環;72 ℃10 min。PCR擴增結束,取5 μL擴增產物于l%瓊脂糖凝膠,100 V電泳20 min,用UVP凝膠成像系統觀察并成像分析。
1.2.7 膠回收
采用北京百泰克生物技術有限公司生產的瓊脂糖回收試劑盒回收目的基因片段,回收方法按試劑盒提供的方法。
1.2.8 葉爾羌高原鰍降落式 PCR基因片段與pMD18-T載體連接
回收的RT-PCR產物與pMD18-T載體于1.5 mL離心管中。于16℃連接過夜。
1.2.9 連接產物的轉化
從-70℃取出感受態克隆細胞DH5α后放置冰水中5 min使之融化,加入連接產物,混勻。冰浴30 min;42℃熱沖擊90 s,冰浴2 min;加入LB液體培養基500 μL,37 ℃水浴10 min,后于37 ℃、200 r/min振蕩培養50 min。取100 μL轉化菌混勻后涂LB Amp+(50 mg/mL)平板,將Amp+板置于37℃培養箱培養12 h。
1.2.10 轉化產物的培養
將轉化的單菌落接種于3 mL LB液體培養基(Amp+50 mg/ml)中,于37℃、160 r/min搖床中培養10 h。
1.2.11 轉化菌的PCR鑒定
反應條件:90℃預變性5 min后進行循環,包括95 ℃、50 s,55 ℃、50 s,72 ℃、50 s,30 個循環后,72℃延伸10 min。
2 結果與分析:
獲得的總RNA中,28S和18S帶型整齊,基本不拖尾,而且28S rRNA帶的亮度明顯高于18S rRNA,無降解,說明RNA完整性好(圖1)。經過降落式PCR目的基因在387 bp,大小與預期的結果基本一致(圖2),而普通PCR并沒有特異性條帶(圖3)。測序后在與加州鱸魚、小口黑鱸抗菌肽Hepcidin同源性達90%以上,在抗菌肽Hepcidin成熟肽有八個半胱氨酸,與已報道的抗菌肽一致(圖4)。通過MEGA4.1構建進化樹,可以看出葉爾羌高原鰍與黑棘鯛在抗菌肽Hepcidin基因親緣關系最近(圖5)。

圖1 葉爾羌高原鰍肝胰臟總RNA
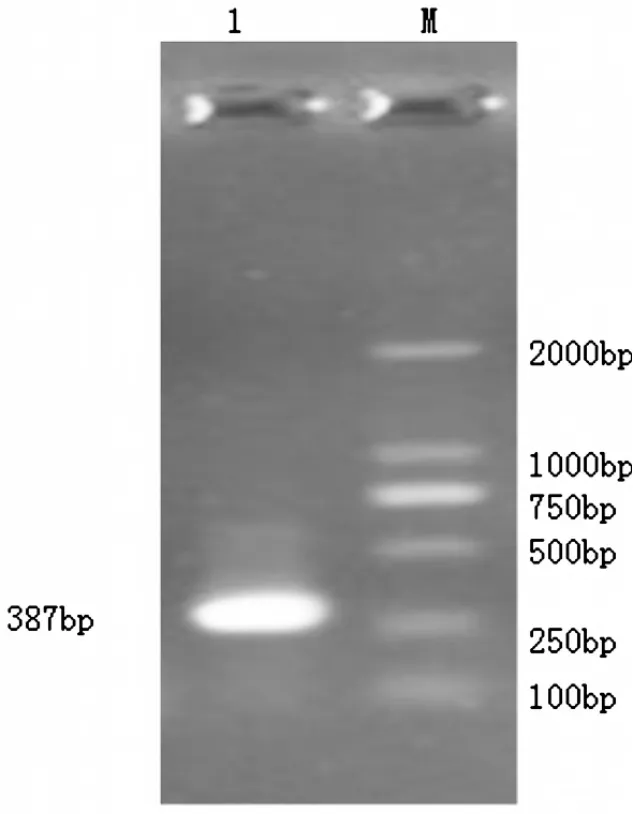
圖2 降落式PCR擴增抗菌肽Hepcidin基因
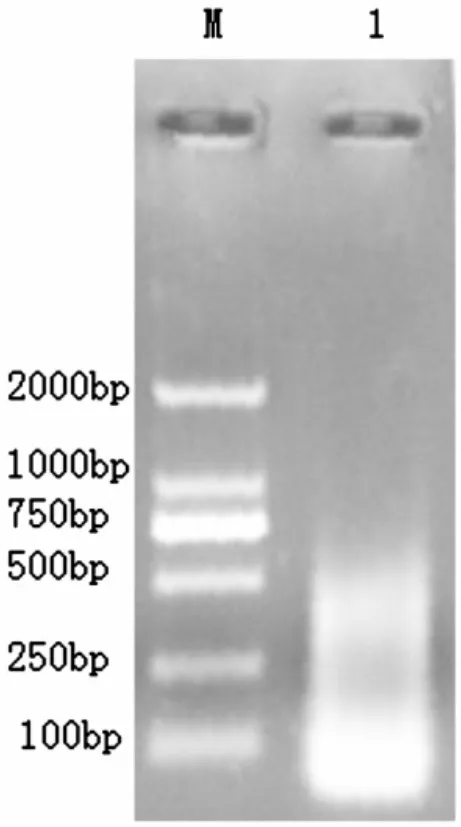
圖3 普通式PCR擴增抗菌肽Hepcidin基因

圖4 葉爾羌高原鰍Hepeidin抗菌肽序列及其編碼的氨基酸序列
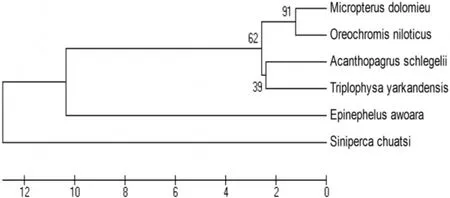
圖5 葉爾羌高原鰍Hepeidin抗菌肽基因進化樹
3 討論
抗菌肽Hepcidi是一類由基因編碼的小分子多肽,是機體免疫防衛系統的重要組成部分,生物體產生的對抗外源性病原體侵襲致病作用的防御性肽類活性物質。抗菌肽分子量小,活性強,功能廣泛。其殺菌過程為抗菌肽分子α-螺旋上的正電荷與細菌質膜磷脂分子上的負電荷在靜電力作用下,相互吸引而靠近;然后借助于分子中N端與C端間的柔性,抗菌肽分子中的疏水端插入質膜中;隨后,α-螺旋也插入質膜中,這樣就破壞了脂質雙分子層原有的有序結構,由于β-折疊的兩親性使抗菌肽分子通過膜內分子間的位移而相互聚集在一起,從而在膜上形成離子通道,細菌最終不能保持正常滲透壓而致死[7,8]。
魚類抗菌肽基因工程方面的報道主要集中在對hepcidin的研究,肝臟是Hepcidin的主要合成場所,脊髓、心臟、肺也有少量的表達,但睪丸、卵巢、前列腺、膀胱、結腸和小腸幾乎沒有表達[9,10]。
本試驗將退火溫度范圍設定為63℃至55℃,主要是為了提高特異性引物與非特異性在該循環范圍內出現的明顯程度。對于低于退火溫度的范圍也會有非特異性,但不明顯[11]。不過與特異性擴增的目的基因相比是微不足道。本實驗經過普通PCR擴增Hepcidin基因重復多次末見特異性條帶,而利用TD-PCR法成功地擴增出了該基因,大小與預期結果及前人的論述一致[12]。
從抗菌肽Hepcidin基因來看普通PCR和TDPCR,前者的優點是:程序省時,簡單,一般的PCR儀都可完成,缺點是:退火溫度不適當時容易出現假陽性或結果不明顯;而TD-PCR程序雖然復雜,但能非常有效地降低或避免假陽性,且在不理想的工作條件下也可擴增出特異性產物,可用于普通PCR難以擴增的基因片段[13]。
通過GenBank數據庫分析發現葉爾羌高原鰍抗菌肽hepcidin基因與黑棘鯛、小口黑鱸同源性達90%,在進化樹可以看出黑棘鯛親緣性最近。從而證明了,降落式PCR可用于普通PCR難以擴增的基因片段。
[1]朱松泉.中國條鰍志[M].南京:江蘇科學技術出版社,1989,129 -131.
[2]中國科學院動物研究所,中國科學院新疆生物土壤沙漠研究所,新疆維吾爾自治區水產局.新疆魚類志[M].烏魯木齊:新疆人民出版社.1979:49-50.
[3]Michael Zasloff.Antimicrobial peptides of multicellular organisms[J].Nature,2002,415(24):389 -395.
[4]Ivana De Domenico,Elizabeta Nemeth,Jenifer M.Nelson,et al.The Hepcidin - Binding Site on Ferroportin Is Evolutionarily Conserved[J].Cell Metabolism ,2008,8(2):146-156.
[5]徐麗君,潘輝,賀艷艷,等.葉爾羌高原鰍肝胰臟總RNA提取方法的比較[J].塔里木大學學報,2010,22(4):18-21.
[6]J.薩姆布魯克,E.F.弗里奇,T.曼尼阿蒂斯.分子克隆實驗指南(第二版)[M].北京:科學出版社,1992.
[7]Chanprapa Imjongjirak,Piti Amparyup,Anchalee Tassanakajon.Molecular cloning,genomic organization and antibacterial activity of a second isoform of antilipopolysaccharide factor(ALF)from the mud crab[J].Scylla paramamosain Fish or Shellfish Immunology,2011,30(1):58-66.
[8]Margitta Dathe,Heike Nikolenko,Jana Meyer,et al.Optimization of the antimicrobial activity of magainin peptides by modification of charge [J].FEBS Letters,2001,501(2 -3):146 -150.
[9]Hunter H N,Fulton D B,Ganz T,et a1.The solution structure of human hepcidin,a peptide hormone with antimicmbial activity that is involved in iron uptake and hereditary hemochromatosis[J].BIO Chem,2002,277(40):37597-37603.
[10]Weinstein D A,Roy C N,Fleming M D,et a1.Inappropriate expression of hepcidin is associated with iron re-fractory anemia:implication for the anemia of chronic disease[J].Blood,2002,100(10):3776 -3781.
[11]張艷萍,郭大瑋,馬莉莉.降落PCR擴增人類常染色體 STR D15S128[J].中國現代醫生,2010,48(2):38-39.
[12]Yin ZhiXin,He Wei,Chen WeiJian,et al.Cloning,expression and antimicrobial activity of an antimicrobial peptide,epinecidin-1,from the orange- spotted grouper,Epinephelus coioides[J].Aquaculture,2006,253(1-4):204-211.
[13]Piraee M,Ving L C.Use of degenerate primers and touchdown PCR to amplify a halogenase gene flagment from Streptomyces venezuelae ISP5230[J].Journal of Industrial Microbiology & Biotechnology,2002,29(1):1-5.
