構象穩定的破傷風毒素Hc片段突變體 (HcM) 的制備及其免疫原性分析
2011-09-29 07:26:12于蕊侯利華劉樹玲于長明張曉艷劉穎陳薇
生物工程學報 2011年2期
關鍵詞:小鼠
于蕊,侯利華,劉樹玲,于長明,張曉艷,劉穎,陳薇
軍事醫學科學院微生物流行病研究所 病原微生物生物安全國家重點實驗室,北京 100071
構象穩定的破傷風毒素Hc片段突變體 (HcM) 的制備及其免疫原性分析
于蕊,侯利華,劉樹玲,于長明,張曉艷,劉穎,陳薇
軍事醫學科學院微生物流行病研究所 病原微生物生物安全國家重點實驗室,北京 100071
破傷風是由破傷風桿菌侵入人體傷口、生長繁殖、產生毒素而引起的一種急性特異性感染,其死亡率高,嚴重危害人民生命健康。研究證實破傷風毒素重鏈 C端 (Hc) 具有與毒素受體結合的活性,完全保留了全分子的免疫原性,有望開發成為新的基因工程破傷風亞單位疫苗以替換傳統的甲醛滅活類毒素疫苗。由于野生型Hc蛋白 (HcW) 易形成分子間及分子內二硫鍵,且各構象分子之間易發生不穩定的轉換,為疫苗的生產工藝帶來困難,因此,通過將破傷風HcW蛋白的869位半胱氨酸突變為丙氨酸,構建構象穩定的破傷風亞單位疫苗突變體HcM,對HcM進行表達、發酵和純化,研究其在不同條件下的穩定性,比較其與受體GT1b親和力的變化及免疫原性。實驗結果表明HcM在大腸桿菌中獲得了高效可溶性表達,通過QXL柱、phenyl-Hs柱和source30Q柱三步純化,可獲得純度95%以上的HcM蛋白。純化后的HcM蛋白在不同保存條件下構象穩定且保持了與破傷風毒素受體GT1b結合的活性。免疫小鼠后,HcM能夠誘導小鼠產生高滴度抗體并能夠保護小鼠抵御1×103LD50劑量的破傷風毒素攻擊。以上結果表明構象穩定的HcM有望成為新的破傷風重組亞單位疫苗候選者,其生產工藝簡單、穩定且保持了與野生型相似的免疫原性,不論是針對日常破傷風治療還是對于國家應對突發事件 (戰爭、地震等災害) 的藥品儲備均具有重大意義。
破傷風,HcM,構象穩定,亞單位疫苗
Abstract:Tetanus is caused by tetanus toxin synthesized by Clostridium tetani. Fragment C (Hc), the 50 kDa carboxy-terminal portion of tetanus toxin, is nontoxic but has receptor protein binding activities, which has been evaluated as a potential new recombinant subunit vaccine to replace the traditional formaldehyde inactivated toxoid vaccine. It is easy for wild Hc (HcW) to form inter- and intra-molecular disulfide bonds and the different conformations changes unstably, which brings difficulties for vaccine production technology. In our study, the Cys 869 of HcW was mutated to Ala and the conformation-stable fragment-C mutant of tetanus toxin (HcM) was constructed. The HcM was expressed, fermented and purified and its stability, receptor binding andimmunogenicity were evaluated. The result showed that the HcM got high-level expression and was purified to >95% of purity. The purified HcM was conformation-stable at different temperature for different time and kept the binding activities with one of its receptor GT1b. Mice given three vaccinations by HcM developed a protective immune response and were 100% protected against an intraperitoneal administration of 1×10350% lethal doses (LD50s) of tetanus neurotoxin. All the results showed that the conformation-stable HcM had potent immunogenicity as a recombinant tetanus vaccine candidate with simple production process and similar immunogenicity with HcW. Whether for routine tetanus therapy or for countries to respond to unexpected events (war,earthquake or other disaster), it is of great significance.
Keywords:tetanus, HcM, conformation-stable, sub-unit vaccine
破傷風是一種嚴重危害人類生命健康的疾病,它是由革蘭氏陽性、厭氧的梭狀芽胞桿菌——破傷風桿菌侵入人體傷口、生長繁殖、產生破傷風毒素引起的一種急性特異性感染,破傷風毒素入侵神經系統可導致人全身性肌肉痙攣,進而全身性衰竭或窒息死亡。一切開放性損傷,均有發生破傷風的可能[1]。破傷風毒素的毒性非常強烈,僅次于肉毒毒素[2],約100 ng的破傷風毒素便可以致人死亡。破傷風在廣大貧困落后的第三世界國家和地區非常嚴重。據估計,世界上每年約有100萬病例發生,死亡率在50%左右。在發展中國家,新生兒破傷風死亡率高達90%。因此各國均致力于尋找高效、安全的破傷風預防藥物[3]。
破傷風毒素全長1 315個氨基酸,分子量為150 kDa,共由A、B、C三部分組成,每部分分子量均為50 kDa。C片段是重鏈的C 端,具有結合神經細胞的作用[4]。B片段是重鏈的 N端,具有導入作用[5]。A片段是輕鏈,有蛋白酶活性,可抑制神經傳遞物質的釋放,具有麻痹作用[6]。傳統的類毒素疫苗是破傷風毒素經甲醛脫毒、鋁佐劑吸附精制而成的,雖然其免疫效果較好,但仍存在一些問題:接種類毒素后有一定的副反應發生;破傷風梭菌可形成芽胞形式,毒性高,生產疫苗有一定危險性;甲醛處理類毒素容易造成污染;化學處理后的類毒素可能發生毒性逆轉;而且重要的是免疫期短,隨著年齡的增長,對破傷風免疫力下降,兒童需要4~6年加強免疫,成人需要每10年加強一次免疫。因此,現有的類毒素疫苗還需進一步改進和發展,而開發新型的基因工程疫苗是方向之一。實驗已證實用完整的毒素分子進行免疫并不是必需的[7]。破傷風天然C 片段 (HcW) 保留了完整毒素與神經節苷脂結合等許多性質[8],免疫效價與毒素相當,無毒性,過敏原性低,是將來發展亞單位疫苗和基因工程疫苗的候選者[9-10],已被用于研究亞單位疫苗、細菌和病毒載體疫苗、粘膜疫苗等很多方面[11]。
野生型HcW分子內含有4個半胱氨酸,極易形成分子間及分子內二硫鍵,導致不同形式的單體及多聚體出現,且這種不同構象形式可隨著蛋白濃度、環境溫度及緩沖液條件的不同而發生相互轉換[12],這在疫苗的生產工藝上造成了一定的困難。為了解決這個問題,我們將HcW分子中的一個關鍵半胱氨酸進行了突變,構建了 Hc突變體HcM,該突變體為構象穩定的單體,簡化了整個疫苗的生產工藝并能夠保證各批次疫苗之間的穩定性。我們對改造后的HcM進行了表達、發酵和純化,并研究了其與受體的結合活性、穩定性、免疫原性等,探討其作為人用破傷風疫苗的可行性,為制備構象穩定的破傷風亞單位疫苗奠定基礎。
通過以上研究,我們希望能夠通過研究制備構象穩定的破傷風亞單位疫苗,為破傷風的預防提供一條更加安全、有效、簡便的道路,其最終目的是希望能夠替代目前使用的存在很多毒副作用及生產繁瑣生產成本高的類毒素疫苗,這對日常破傷風治療和國家應對突發事件 (戰爭、地震等災害) 的藥品儲備均具有重大意義。
1 材料與方法
1.1 材料
Q Sepharose XL柱、Phenyl-HS柱和source30Q柱購自通用電氣 (中國) 醫療集團。DNA 核酸內切酶 EcoRⅠ、XhoⅠ和 T4 DNA連接酶購自寶生物工程 (大連) 有限公司。Isopropylthio-D-galactoside(IPTG) 和神經節苷酯GT1b購自Sigma。Al(OH)3佐劑購自 Brenntag Biosector (丹麥)公司。表達載體pET-32a(+) 和其宿主菌 E. coli BL21 (DE3) 購自Novagen。野生型HcW基因片段為本實驗室全合成后保存。HcW蛋白為本實驗室表達純化后保存。小鼠抗破傷風類毒素多抗血清為本實驗室制備保存。HRP標記的羊抗鼠 IgG購自中杉金橋北京有限公司。BALB/c小鼠購自軍事醫學科學院實驗動物中心。破傷風毒素和甲醛滅活的破傷風類毒素均為國家藥品生物制品檢定所贈送。
1.2 方法
1.2.1 HcM的原核可溶性高表達
選用本實驗室之前全合成的 HcW (865-1351)基因片段為模板,上游引物aaaaacttgatgcatgggtcgacaacgaagaag-3′ (下劃線部分為EcoRⅠ酶切位點) 和下游引物 MR:5′-ccgctcgag tcattagtcgttggtccaaccttcatc-3′ (下劃線部分為 XhoⅠ酶切位點),采用probest DNA聚合酶,致突變PCR擴增將原 HcW片段中的 Cys869 (tgt) 突變為Ala869(gca),從而獲得完整的HcM DNA片段。HcM片段回收、酶切后連接入 EcoRⅠ和 XhoⅠ雙酶切后的pET32a(+) 載體中,轉化后挑取陽性克隆送測序。將測序正確的HcM表達質粒轉化E. coli BL21 (DE3),挑取單克隆至5 mL LB液體培養基中,37 ℃、220 r/min培養至 OD600≈0.6時,加入終濃度為0.2 mmol/L的IPTG,28 ℃、220 r/min繼續誘導表達4 h。離心收獲菌體沉淀,PBS重懸后超聲碎菌,離心取上清,進行 12% SDS-PAGE蛋白電泳以檢測目的蛋白的表達。
1.2.2 無標簽HcM的發酵及純化
表達HcM蛋白的種子液1 L,轉接入30 L發酵罐中,培養至OD600≈0.6時,加入終濃度為0.2 mmol/L的 IPTG,28 ℃、300 r/min繼續培養 4 h。10 000×g、4 ℃離心收集菌體,用20 mmol/L Tris-HCl緩沖液(pH 8.5) 重懸后勻漿碎菌。20 000×g、4 ℃離心收集上清。上清液過Q Sepharose XL柱進行陰離子交換,用20 mmol/L Tris-HCl緩沖液 (pH 8.5) 平衡后,用含0~0.5 mol/L NaCl的Tris-HCl緩沖液進行梯度洗脫。含有目的蛋白的洗脫液加入終濃度為0.8 mol/L的 (NH4)2SO4后,過苯基疏水phenyl-HS柱進行下一步純化。目的蛋白用含濃度0.8~0 mol/L的 (NH4)2SO4的 Tris-HCl (pH 8.5) 緩沖液進行梯度洗脫。含有目的蛋白的洗脫液用脫鹽柱置換緩沖液為20 mmol/L Tris-HCl (pH 8.5) 后,過source30Q柱進行進一步的陰離子交換,目的蛋白用含0~0.5 mol/L NaCl的20 mmol/L Tris-HCl緩沖液 (pH 8.5) 進行梯度洗脫,收集產物用SDS-PAGE蛋白電泳確定蛋白純度。
1.2.3 HcM和HcW構象穩定性比較
將純化后的HcM及HcW蛋白分別于4 ℃、?20 ℃、?80 ℃保存0 d、2 d、7 d和2個月,采用非還原SDS-PAGE檢測蛋白的構象穩定性。
1.2.4 HcM和HcW與神經節苷酯GT1b的結合
神經節苷酯 GT1b用甲醇稀釋至 5 μg/mL,100 μL/孔包被96孔酶聯板,室溫放置過夜至甲醇完全揮發,PBST洗滌5 min,重復4次。將HcM和HcW分別用PBST從20 μg/mL進行倍比稀釋,100 μL/孔加入96孔板中,37 ℃孵育1 h。PBST洗滌5 min,重復4次后加入1∶800稀釋的小鼠抗破傷風類毒素多抗血清,100 μL/孔,37 ℃繼續孵育1 h。PBST洗滌5 min,重復4次后加入1∶4 000稀釋的HRP標記山羊抗小鼠IgG二抗,37 ℃反應30~40 min。加入 TMB 顯色液,50 μL/孔,顯色后用 2 mol/L H2SO4終止反應,酶標儀450 nm測定光吸收值。
1.2.5 HcM和HcW的免疫原性比較
20 μg/mL的HcM和HcW蛋白加入終濃度為1.5 mg/mL的Al(OH)3佐劑制成疫苗懸液,免疫6~8周齡雌性 Balb/c小鼠,0.5 mL/只進行腹部皮下注射,6只/組,同時設Al(OH)3佐劑陰性對照組,每2周免疫1次,共免疫3次,每次免疫后2周、下一次免疫前取血檢測血清中的抗體滴度;抗體滴度的測定采用ELISA的方法,破傷風類毒素1 μg/mL,100 μL/孔包被96孔酶聯板,4 ℃包被過夜。PBST洗滌4次,5 min/次。將抗血清用PBST從1∶100依次倍比稀釋后,加入酶聯板中,37 ℃反應1 h,同時設PBS免疫后的血清為對照。PBST洗滌4次,5 min/次。加入 HRP標記山羊抗小鼠 IgG二抗,37 ℃反應30~40 min。加入TMB顯色液,50 μL/孔,顯色后用2 mol/L H2SO4終止,酶標儀450 nm測定光吸收值。以加入陰性對照血清孔的顯色值為對照,以OD450大于0.1且高于陰性孔2倍為陽性稀釋滴度。1.2.6 HcM對小鼠的攻毒保護性
將破傷風毒素用硼酸鹽緩沖液稀釋,腹腔注射HcM和HcW免疫3次后的小鼠,每只小鼠注射0.5 mL,含1×103LD50的破傷風毒素,每組6只小鼠,同時設陰性對照組。攻毒后每天觀察并記錄小鼠的發病及存活情況。
2 結果與分析
2.1 HcM的原核可溶性高表達
PCR擴增得到的 HcM 片段經酶切后連接入原核表達載體pET32a(+) 中,在大腸桿菌BL21 (DE3)中獲得了高效可溶性表達 (圖 1),目的蛋白分子量為50 kDa左右,主要以可溶性的形式存在于碎菌上清中,約占碎菌上清總蛋白的40%。
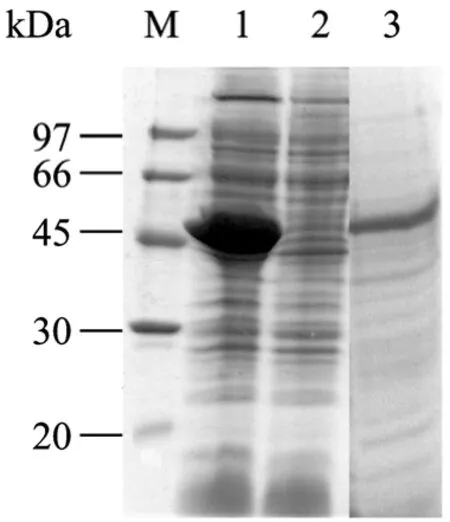
圖 1 HcM 在 BL21(DE3) 表達碎菌上清及沉淀中的SDS-PAGE蛋白電泳圖Fig. 1 SDS-PAGE detection of HcM expressed in the ultrasonic supernatants of BL21(DE3). M: protein marker; 1:supernatant of pET32a(+)-HcM; 2: supernatant of pET32a(+);3: precipitation of pET32a(+)-HcM.
2.2 無標簽HcM的發酵與純化
HcM蛋白在30 L發酵罐中成功進行了發酵,發酵罐中pH控制在7.0±0.5,溶氧控制在30%~80%,通氣量3~6 L/min,罐壓控制在0~0.05 MPa,28 ℃誘導表達4 h后,收獲菌體沉淀,經勻漿后離心收集表達上清,依次過Q Sepharose XL柱、Phenyl-HS柱和 source30Q柱進行純化,經過三步純化后,無標簽的目的蛋白 Hc得到了很好地純化,純度可達95%以上 (圖2),得率在300 mg/L以上,目的蛋白最終保存在PBS緩沖液中。
圖2 三步純化后獲得的HcM的SDS-PAGE蛋白電泳圖Fig. 2 SDS-PAGE detection of purified HcM after 3 steps purification. M: protein marker; 1,2: purified HcM.
2.3 HcM的構象穩定性
將純化后的HcM和HcW分別在4 ℃、?20 ℃、?80 ℃放置不同時間,觀察它們的構象穩定性。實驗結果表明HcW存在多聚體及2種構象形式的單體——氧化型和還原型,新鮮純化后的HcW主要以還原型為主,在4 ℃放置2 d后,HcW由還原型向氧化型轉變,2種單體構象形式約各占一半;4 ℃放置7 d及2個月后,HcW主要以氧化型單體存在且多聚體也有明顯增加。而HcM在4 ℃放置2個月以上仍能保持構象的穩定 (圖 3)。HcW 和 HcM 在?20 ℃、?80 ℃放置不同時間后各構象形式均基本穩定 (數據未顯示)。
2.4 HcM與受體神經節苷酯GT1b的結合活性
采用ELISA的方法檢測了HcM及HcW與破傷風毒素受體之一 GT1b的結合活性,結果表明 (圖4),與HcW相比,HcM與GT1b的結合能力較弱,這也意味著HcM引起體內病理學反應的幾率降低,從而可以降低副作用,更加適合用作人用疫苗。
2.5 HcM的免疫原性分析
采用Al(OH)3為佐劑,HcM和HcW為蛋白疫苗分別免疫小鼠3次,每2周免疫1次,從每次免疫后測得的小鼠抗破傷風類毒素滴度 (圖 5) 可以看出,第1次免疫后 2周,2組小鼠抗體的平均滴度均在1∶500左右;第2次免疫后,兩組小鼠抗體的平均滴度均高于 1∶10 000,且 HcM 組略高于HcW組;第3次免疫后,2組小鼠抗體的滴度均為 1∶100 000左右,且2組小鼠的抗體滴度之間無明顯差別。
圖3 HcW和HcM在4 ℃放置不同時間的構象穩定性Fig. 3 The conformational stability of HcW and HcM at 4 °C for different time. A: 4 °C for 0 d; B: 4 °C for 2 d; C: 4 °C for 7 d; D:4 °C for 2 months. M: protein marker.
圖4 不同濃度的HcW和HcM與神經節苷酯GT1b的結合活性Fig. 4 The binding of GT1b with HcW and HcM in different concentration.
圖5 HcW和HcM免疫小鼠后不同時間測得的抗破傷風類毒素抗體滴度Fig. 5 The anti-tetanus toxoid antibody titers of mice after HcW and HcM immunization.
表1 HcW和HcM作為亞單位疫苗對小鼠的體內保護作用Table 1 In vivo protection of HcW and HcM used as sub-unit vaccine
2.6 HcM對小鼠的攻毒保護性
用含1×103LD50的破傷風毒素攻擊免疫3次后的小鼠,觀察小鼠的存活情況,從實驗結果 (表 1)可以看出,10 μg的 HcM 可以完全保護小鼠抵御1×103LD50的破傷風毒素攻擊,其保護效果同HcW相當。
3 討論
破傷風毒素的毒性非常強烈,僅次于肉毒毒素,約100 ng的破傷風毒素便可以致人死亡,因此極有可能被用于生物戰劑。1997年召開的生物武器公約締約國政府專家特設小組第七次會議確定的生物戰劑標準與戰劑病原體清單中,破傷風毒素被列為用于攻擊人的毒素。安全、有效的破傷風疫苗對于部隊的平、戰時均具有特殊意義,在進行野外作戰、執行維和任務及抗震救災時可作為藥品儲備及安全保障。一切開放性的傷口均由發生破傷風的可能,隨著我國飼養寵物的家庭越來越多,感染破傷風的風險和幾率也在逐漸擴大,基因工程破傷風亞單位疫苗的研制成功,將使廣大民眾能接種安全有效的新型疫苗,減少破傷風發病率,具有良好的應用前景。
目前國內外均沒有重組破傷風疫苗臨床前或臨床研究的報道,原因是表達純化的Hc蛋白存在表達量較低、易形成包涵體且免疫原性低等問題。本實驗室已經在大腸桿菌中成功獲得了野生型 HcW 蛋白的可溶性高表達,其免疫原性與類毒素相當,但由于破傷風毒素HcW片段在869、1077、1 093和1 301位存在4個半胱氨酸cys,其中869位和1 093位的cys易形成分子內二硫鍵,其余的cys可形成分子間二硫鍵[12],因此其構象極其不穩定,這給疫苗后續的生產工藝帶來了一定的困難,因此,本研究通過將Hc片段869位關鍵的cys突變為Ala,使其形成條帶均一、構象穩定的蛋白HcW,并且對其蛋白穩定性、受體結合能力及免疫原性進行了分析,實驗結果表明突變后的蛋白保持了單一的單體構象,在4 ℃放置2個月以上仍能保持構象的穩定;盡管與 HcW 相比,HcM與受體神經節苷酯 GT1b的結合活性略有下降,但這也意味著HcM在體內引起的病理反應也會相應減少,從而減少疫苗的毒副作用。但突變后的HcW仍能夠誘導小鼠產生高滴度的抗破傷風類毒素抗體,并能保護小鼠抵御至少1×103LD50的破傷風毒素的攻擊。
構象穩定的破傷風HcW蛋白具有開發成為新型破傷風重組亞單位疫苗的潛力,其成功研制可為破傷風的預防提供一條更加安全、有效、簡便的道路。
REFERENCES
[1] Mallick IH, Winslet MC. A review of the epidemiology,pathogenesis and management of tetanus. Int J surg, 2004,2(2): 109?112.
[2] Lacy DB, Tepp W, Cohen AC, et al. Crystal structure of botulinum neurotoxin type A and implications for toxicity.Nature Struct Biol, 1998, 5(10): 898?902.
[3] Quinn HE, McIntyre PB. Tetanus in the elderly –An important preventable disease in Australia. Vaccine, 2007,25(7): 1304?1309.
[4] Bizzini B, Stoeckel K, SchwabM. An antigenic polypeptide fragment isolated from tetanus toxin:chemical characterization, binding to gangliosides and retrograde axonal transport in various neuron systems. J Neurochem, 1977, 28(3): 592?542.
[5] Matsuda M, Lei DL, Sugimoto N, et al. Isolation,purification, and characterization of fragment B, the NH22-terminal half of the heavy chain of tetanus toxin.Infect Immun, 1989, 57(11): 3588?3593.
[6] Schiavo G, Poulain B, Rossetto O, et al. Tetanus toxin is a zinc protein and its inhibition of neurotiansmitter release and protease activity depend on zinc. EMBO J, 1992,11(10): 3577?3583.
[7] Lafaye P, Nato F, Mazié JC, et al. Similar binding properties for a neutralizing anti-tetanus toxoid human monoclonal antibody and its bacterially expressed Fab.Res Immunol, 1995, 146(6): 373?382.
[8] Rummel A, Bade S, Alves J, et al. Two carbohydrate binding sites in the Hcc-domain of tetanus neurotoxin arerequired for toxicity. J Mol Biol, 2003, 326(3): 835?847.
[9] Halpern JL, Loftus A. Characterization of the receptor-binding domain of tetanus toxin. J Biol Chem,1993, 268(15): 11188?11192.
[10] Herreros J, Lalli G, Schiavo G. C-terminal half of tetanus toxin fragment C is sufficient for neuronal binding and interaction with a putative protein receptor. Biochem J,2000, 347(Pt 1): 199?204.
[11] Wells JM, Wilson PW, Norton PM, et al. Lactococcus lactis: high-level expression of tetanus toxin fragment C and protection against lethal challenge. Mol Microbiol,1993, 8(6): 1155?1162.
[12] Qazi O, Bolgiano B, Crane D, et al. The HCfragment of tetanus toxin forms stable, concentration-dependent dimers via an intermolecular disulphide bond. J Mol Biol,2007, 365(1): 123?134.
Production and immunogenicity analysis of conformation-stable fragment-C mutant of tetanus toxin
Rui Yu, Lihua Hou, Shuling Liu, Changming Yu, Xiaoyan Zhang, Ying Liu, and Wei Chen
State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, China
Received: July 8, 2010; Accepted: November 12, 2010
Supported by: National Basic Research Program of China (973 Program) (No. 2006CB504404), Important National Science and Technology Specific Projects (No. 2008ZX10003-010), National Department Public Benefit Research Foundation (No. 200903027).
Corresponding author: Xin’an Jiao. Tel: +86-514-87971803; E-mail: jiao@yzu.edu.cn*These authors contributed equally to this work.
國家重點基礎研究發展計劃 (973計劃) (No. 2006CB504404),國家科技重大專項 (No. 2008ZX10003-010),公益性行業科研專項 (No. 200903027)資助。
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
