新型抗結核分枝桿菌藥物篩選模型的建立
2011-09-25 09:30:26張磊磊由鵬飛姚廣新侯爵張怡軒
微生物學雜志 2011年1期
張磊磊,由鵬飛,2,姚廣新,侯爵,張怡軒,2*
(1.沈陽藥科大學生物制藥教研室,遼寧沈陽110016;2.沈陽藥科大學生物學教學實驗中心,遼寧沈陽110016)
新型抗結核分枝桿菌藥物篩選模型的建立
張磊磊1,由鵬飛1,2,姚廣新1,侯爵1,張怡軒1,2*
(1.沈陽藥科大學生物制藥教研室,遼寧沈陽110016;2.沈陽藥科大學生物學教學實驗中心,遼寧沈陽110016)
為建立基于酶水平和細胞水平的新型抗結核分枝桿菌(Mycobacterium tuberculosis)藥物的篩選模型,以M.tuberculosis H37Rv基因組DNA為模板,PCR特異性擴增異檸檬酸裂解酶(ICL)基因,構建表達載體,在E.coli BL21(DE3)中高效表達,使用Ni2+親和層析柱純化重組ICL,檢測其活性。優化ICL酶反應條件,考察待篩選樣品溶劑對酶活性的影響,建立ICL抑制劑酶水平篩選模型;考察與優化恥垢分枝桿菌(Mycobacterium smegma)在以乙酸鹽為唯一碳源的培養基中的生長狀況,建立基于M.smegma的乙醛酸代謝途徑抑制劑的細胞水平篩選模型;利用上述2種篩選模型對1060種可能具有拮抗活性的微生物代謝樣品進行初篩和復篩,兩者篩選結果正相關性較好。
結核分枝桿菌;異檸檬酸裂解酶;恥垢分枝桿菌;乙醛酸途徑;篩選模型
結核病是由結核分枝桿菌(Mycobacterium tuberculosis)引起的傳染性疾病。WHO報告顯示,全球有超過20億人感染M.tuberculosis,其中約有1/10的人會發展為結核病。2008年全球新增940萬結核病病例,同年180萬人死于結核病。目前多重耐藥結核分枝桿菌(MDR-TB)的出現及M.tuberculosis在患者體內的持留狀態,已經成為結核病預防和治療過程中的重大障礙,因此針對M.tuberculosis在患者體內的持留狀態,尋找新的藥物作用靶點,建立新的藥物篩選模型是開發新型抗結核藥物的關鍵。致病性菌株M.tuberculosis H37Rv全基因組測序工作的完成,為研究提供了大量的藥物靶點[1]。異檸檬酸裂解酶(ICL)作為乙醛酸途徑的關鍵酶,當培養基的碳源受到限制或缺氧時,菌體內糖酵解途徑減弱,乙醛酸循環加強,酶活性上調[2]。Mckinney等[3]的研究表明ICL對M.tuberculosis在巨噬細胞內的持續存活至關重要。乙醛酸途徑存在于原核生物、低等真核生物和植物中,在哺乳動物體內不存在[4],故可將ICL作為特異性極強的藥物靶點,建立新型抗結核分枝桿菌藥物的篩選模型[5]。但是以往的研究僅僅針對體外ICL酶活性進行抑制劑的篩選,對于活性化合物是否具有抑制菌體內ICL酶活性并沒有進一步的篩選方法。由于M.tuberculosis具有極強的傳染性和致病性,故培養M.tuberculosis需要在P2或者P3實驗室進行,實驗危險性大、成本高。本實驗利用無毒的恥垢分枝桿菌(Mycobacterium smegma)替代M.tuberculosis進行研究,考察以乙酸鹽為唯一碳源的M.smegma生長狀況以及乙醛酸途徑的啟動,建立乙醛酸途徑抑制劑細胞水平的篩選模型。本文針對ICL分別建立酶水平和細胞水平的篩選模型,即首先克隆表達M.tuberculosis H37Rv的ICL,建立酶水平抑制劑篩選模型;又以乙酸鹽為唯一碳源,建立M.smegma乙醛酸代謝途徑抑制劑的細胞水平篩選模型。利用上述2種模型對本實驗室的微生物發酵樣品進行篩選,正相關性達到90%以上。
1 材料與方法
1.1 材料
1.1.1 實驗材料M.tuberculosis H37Rv icl基因由復旦大學趙國屏教授惠贈;E.coli BL21(DE3) plysS、E.coli DH-5α、M.smegma(ATCC607)由本實驗室保藏;質粒pET-28b(+)由本實驗室保藏;微生物發酵樣品1060種(本室分離并保存)。
1.1.2 試劑T4 DNA-ligase、限制性內切酶NdeⅠ、HindⅢ,pMD18-T Vector,Ex Taq Polymerase均購自TAKARA公司;3S柱離心式DNA小量快速純化試劑盒、3S柱離心式質粒小量快速抽提試劑盒、BBST NTA-Ni Resin購自上海申能博彩生物科技有限公司;考馬斯亮蘭蛋白測定試劑盒購自南京建成生物工程研究所;MarkerⅣ、Protein Molecular Weight Marker購自天根生化科技(北京)有限公司。
1.2 方法
1.2.1 M.tuberculosis H37Rv ICL基因的克隆
以M.tuberculosis H37Rv icl基因為模版,使用Primer(S):5'-GGGTTT CATATG TCTGTCGTCGGCAC-3'(下劃線部分為NdeⅠ酶切位點)、Primer (A):5'-CCC AAGCTT CTAGTGGAACTGGCCC-3' (下劃線為部分為HindⅢ酶切位點)為模板進行聚合酶鏈反應(PCR),擴增目的基因。PCR反應條件為95℃變性10 min、95℃變性30 s、55℃退火30 s、72℃延伸90 s、72℃延伸10 min,共進行30個循環。擴增產物經瓊脂糖凝膠電泳檢測無誤后回收純化。將純化的PCR產物與pMD18-T Vector進行連接,轉化入E.coli DH-5α中。重組克隆載體命名pMD18-ICL。抽取克隆載體經NdeⅠ、HindⅢ雙酶切后純化ICL片段并與pET-28b(+)經NdeⅠ、HindⅢ酶切后的純化產物進行連接,轉化入E.coli DH-5α中。挑取陽性克隆,提取質粒,命名為pET-28b-ICL。
1.2.2 重組ICL的表達與純化將重組質粒pET-28b-ICL轉化入表達菌株E.coli BL21 (DE3)plysS,LB/Kan平板篩選陽性克隆,挑取單菌落于5 mL LB培養基(Kan 50 g/mL)中,37℃220 r/min培養16 h,取3 mL菌液轉種至50 mL LB培養液(Kan 50 μg/mL)中,37℃220 r/ min培養2~3 h,加入IPTG(終濃度0.25 mmol/ L)25℃,220 r/min培養6 h,離心收集,加入1/ 20菌體培養液體積的NTA-0 Buffer(20 mmol/L Tris-HCl pH 7.9、0.5 mmol/L NaCl、10%Glycerol)和PMSF(終濃度為1 mmol/L)懸浮菌體,置于冰上超聲破碎(40%功率,超聲1 s,間隔1 s,超聲總時間為5 min),分別收集沉淀和上清,并設立E.coli BL21(DE3)plysS與含有pET-28b的E.coli BL21(DE3)plysS為對照,采用十二烷基硫酸鈉聚丙烯酰胺凝膠(SDS-PAGE)電泳檢驗重組ICL的表達狀況。用4倍柱體積的超聲上清液上樣NTA-Ni Resin,以含20、40、60、80、200 mmol/L咪唑的NTA buffer對目的蛋白進行洗脫,使用SDS-PAGE來判斷目的蛋白在洗脫液中的分布狀況。使用考馬斯亮蘭染色法測定ICL蛋白濃度。
1.2.3 重組ICL活性檢測及動力學參數測定
ICL能催化異檸檬酸分解為乙醛酸和琥珀酸,乙醛酸與苯肼反應,生成物苯腙在324 nm處有紫外吸收,以此測定重組ICL的活性。參照文獻[6]建立ICL活性檢測的基礎體系:MgCl2·6H2O 7.5 mmol/L,鹽酸苯肼5 mmol/L,DL-半胱氨酸3 mmol/L,Tris-HCl 50 mmol/L,DL-異檸檬酸三鈉鹽2.5 mmol/L。分別考察pH、Mg2+濃度和DL-半胱氨酸濃度對酶活性的影響,以確定最佳酶活檢測體系,并測定重組ICL比活力。在此條件下,以0.03~1 mmol/L的異檸檬酸為底物,測定ICL的反應速度。根據酶催化底物的[S]-1-υ-1關系繪制曲線。通過Lineweave-Bure方程計算重組ICL米氏常數(Km)。
1.2.4 ICL抑制劑酶水平篩選模型的建立由于所篩選樣品為微生物發酵液乙酸乙酯萃取物,在酶促反應體系中溶解度較差,嚴重干擾篩選結果。因此實驗過程中選用DMSO和其他表面活性劑共同溶解樣品。實驗中考察了終濃度(體積比)為1%、2%、4%、6%、8%、10%和20%的DMSO;終濃度(體積比)為0.1%、0.25%、0.5%、1%、2%的Tween-20;終濃度(質量與體積比)為0.1%、0.25%、0.5%、1%的SDS對ICL活性的影響。由此確定篩選體系中樣品溶劑的最佳加入量,結合ICL最適反應條件建立ICL抑制劑篩選模型。
1.2.5 乙醛酸途徑抑制劑細胞水平篩選模型的建立采用改良的Dubos培養基(Apsaragine 2 g/ L,Na2HPO42.5 g/L,KH2PO41g/L,MgSO4·7H2O 10 mg/L,枸櫞酸鐵氨50 mg/L,CaCl20.5 mg/L,ZnSO40.1 mg/L,CuSO40.1 mg/L,Tween-800.05%,NaAc 10 mmol/L,)培養M.smegma,使之利用乙酸鹽作為唯一碳源[2],啟動菌體內乙醛酸循環。考察在這種培養基中M.smegma的生長狀態以及菌體內ICL活性,以確定篩選樣品最佳加入時間。由于使用DMSO溶解待篩選樣品,實驗中考察了終濃度(體積比)為1%、2%、3%、4%的DMSO對菌體生長的影響。
2 結果與分析
2.1 ICL基因的克隆與表達
以M.tuberculosis H37Rv基因組DNA為模版進行PCR擴增,以0.7%瓊脂糖凝膠電泳檢測,擴增產物約為1300 bp大小的特異性條帶,與預期結果一致(圖略)。將PCR產物連接入載體pMD18-T Vector,通過藍白斑篩選挑取陽性克隆,經雙酶切獲得2個大小分別為1300 bp和2700 bp的片段,與預期結果一致(圖略)。鑒定后將目的基因片段克隆入表達載體pET-28b(+),重組質粒經雙酶切驗證、PCR驗證、測序驗證正確后命名為pET-28b-ICL。
2.2 重組ICL的表達與純化
將重組質粒pET-28b-ICL轉化E.coli BL21 (DE3)plysS中,誘導表達后超聲破碎菌體,經SDS-PAGE檢測后發現在50 ku處有重組蛋白條帶,并且超聲上清液中的目的蛋白含量遠大于沉淀中的目的蛋白含量(圖1),說明目的蛋白大部分可溶性表達,因此選用超聲上清液進一步純化目的蛋白。
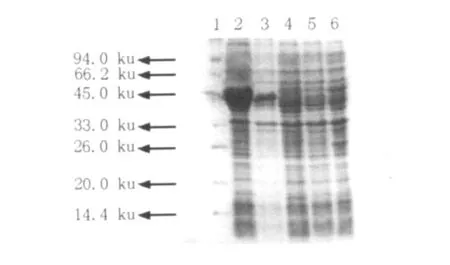
圖1 SDS-PAGE檢測重組ICL表達情況Fig.1Detection of the expression of recombinant ICL by SDS-PAGE 1:Protein Molecular Weight Marker;2:E.coli BL21(pET-28b-ICL)ultrasonic supernatant after induction;3:E.coli BL21(pET-28b-ICL)ultrasonic precipitation after induction;4:E.coli BL21(pET-28b-ICL)non-induced;5:E.coli BL21(pET-28b)after IPTG induction;6:E.coli BL21 afer IPTG induction
洗脫液經SDS-PAGE檢測,結果顯示目的蛋白主要存在于含200 mmol/L咪唑的洗脫緩沖液中(圖2),且達到電泳純。透析除去洗脫液中的咪唑,并經適當濃縮,測得純化后ICL濃度為0.36 mg/mL。
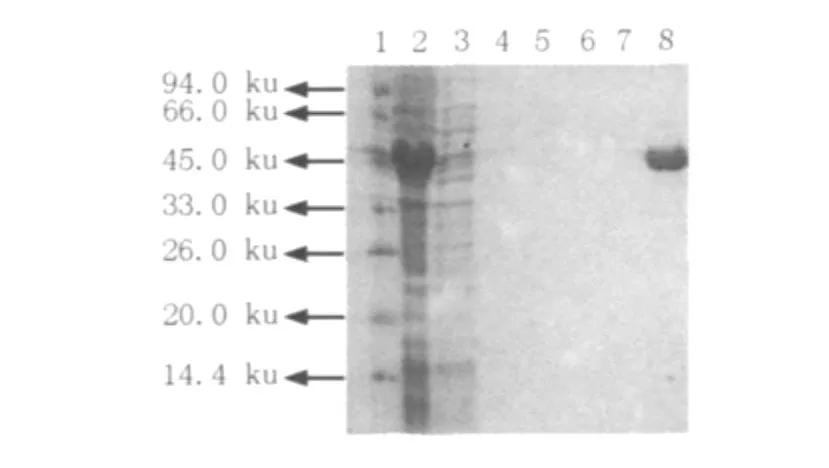
圖2 SDS-PAGE檢測重組ICL純化情況Fig.2Detection of the purification of recombinant ICL by SDS-PAGE 1:Protein Molecular Weight Marker;2:E.coli BL(pET-28b-ICL)ultrasonic supernatant after induction;3~8:Eluent with imidazole(0、20、40、60、80、200 mmol/L)
2.3 重組ICL活性檢測及動力學參數測定
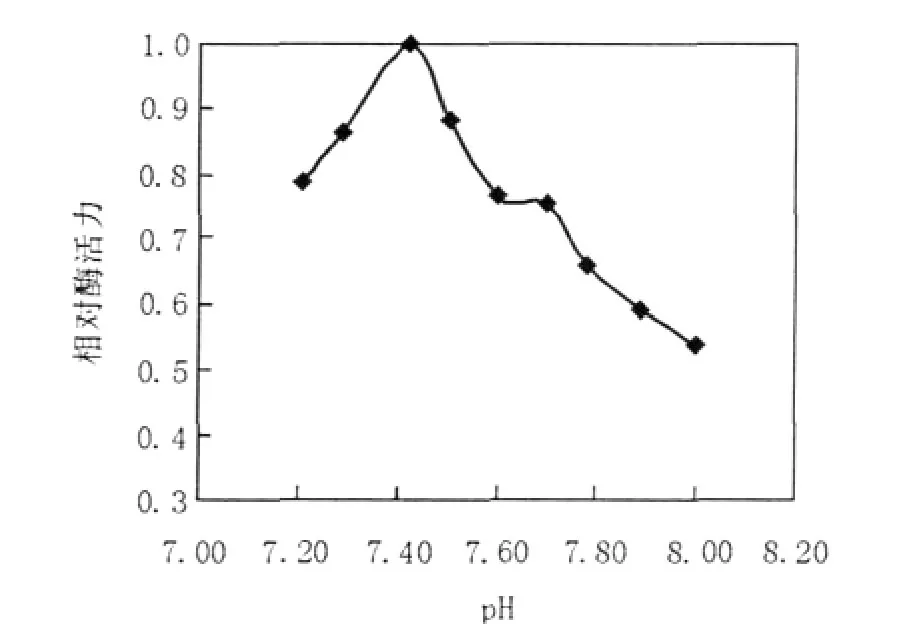
圖3 pH對ICL活力的影響Fig.3The effect of pH on the activity of the recombinant ICL
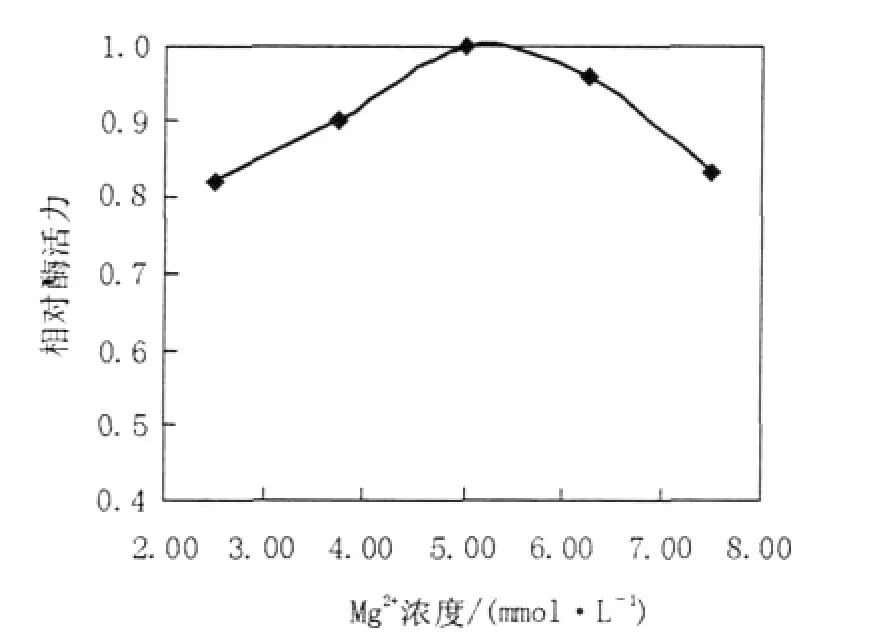
圖4 Mg2+濃度對ICL活性的影響Fig.4The effect of different concentrations Mg2+ on the activity of the recombinant ICL
于37℃,在不同pH值(分別為7.21、7.29、7.42、7.50、7.60、7.70、7.78、7.89、8.00)的100 mmol/L Tris-HCl緩沖液中測定重組ICL的活性,實驗結果顯示ICL最適pH為7.42(圖3);在最適pH條件下調整反應體系中Mg2+濃度(分別為2.5、3.75、5、6.25、7.5 mmol/L),最終確定Mg2+最佳濃度為5 mmol/L(圖4);調整反應體系中DL-半胱氨酸終濃度(分別為3.5、5.0、7.0 mmol/L),最終確定DL-半胱氨酸最適終濃度為5.0 mmol/L (圖5)。因此,最適酶促反應體系:100 mmol/L Tirs-HCl(pH 7.42)50 μL,50 mmol/L MgCl210 μL,5.0 mmol/L DL-半胱氨酸10 μL,0.1 mol/L鹽酸苯肼5 μL,10 mmol/L DL-異檸檬酸三鈉鹽2 μL,0.36 mg/mL ICL 3 μL。在此條件下,對ICL的比活力進行測定,測得純化后ICL的比活力為1.402 μmol/min/mg,其Km值為0.811 mmol/L。
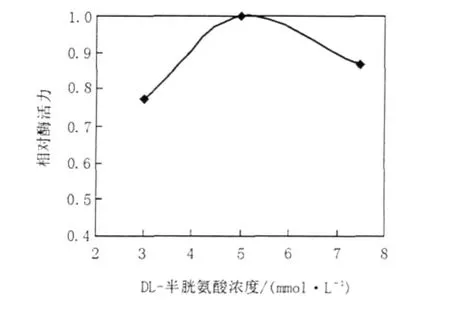
圖5 DL-半胱氨酸濃度對重組ICL活性的影響Fig.5The effect of different concentrations of DL-Cys on the activity of the recombinant ICL
2.4 ICL抑制劑酶水平篩選模型的建立
實驗證明,終濃度(體積比)為1%~20%的DMSO對ICL的活性并無顯著影響,因此可以作為待篩選樣品的溶劑。在0.5%~2%Tween-20對ICL活性的影響呈濃度相關性,濃度越高,ICL活性損失越多。但當Tween-20終濃度(體積比)為0.25%時ICL活性幾乎沒有損失,而且表現出輕微激活作用,因此在篩選體系中加入終濃度為0.25%的Tween-20,使樣品增溶。終濃度(質量與體積比)在0.1%~1%范圍內,SDS顯著抑制ICL活性,損失率達到95%以上,因此不予采用。故最終確定待篩選樣品的溶劑系統為100 μL DMSO與100 μL 2.5%Tween-20的混合液。最終確定ICL酶水平抑制劑篩選體系:Tirs-HCl(pH 7.42)50 mmol/L, MgCl25 mmol/L,DL-半胱氨酸0.5 mmol/L,鹽酸苯肼5 mmol/L,DL-異檸檬酸三鈉鹽0.2 mmol/L,待篩選樣品溶液20 μL,ICL 10.8 μg/mL。
2.5 ICL抑制劑酶水平篩選步驟
取96孔板,每孔加入100 mmol/L Tirs-HCl (pH 7.42)50 μL,50 mmol/L MgCl210 μL,5.0 mmol/L DL-半胱氨酸10 μL,0.1 mol/L鹽酸苯肼5 μL,待篩選樣品溶液20 μL,ICL(0.36 mg/mL) 3 μL。將96孔板置于振蕩器震蕩5min,再置于37℃溫浴15 min,324 nm處檢測,吸光度值記錄為OD1,每孔加入10 mmol/L DL-異檸檬酸三鈉鹽2 μL,振蕩器震蕩5 min,37℃溫浴15 min,振蕩器震蕩5 min,324 nm檢測,吸光度值記錄為OD2,同時設立樣品的溶劑為陰性對照,其吸光度變化值記做Δ空白。樣品抑制率計算公式如下。
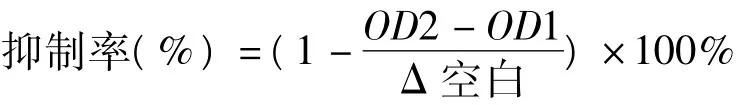
2.6 乙醛酸途徑抑制劑細胞水平篩選模型
實驗結果表明,M.smegma在以乙酸鹽為唯一碳源的培養基中培養13 h后,菌種進入對數生長期,菌液呈均勻渾濁狀,其濁度在OD600顯著增長(圖6),并一直持續到22 h,22 h后菌體結團、有顆粒,不利于菌體濃度的檢測;檢測了對數生長期內15 h和20 h對應的培養液中菌體超聲上清液ICL酶活性,發現其顯著提高(圖7),說明菌體確實處于乙醛酸代謝途徑中,而且ICL酶活性增強,因此選取培養15~20 h進行ICL抑制劑的細胞水平篩選。
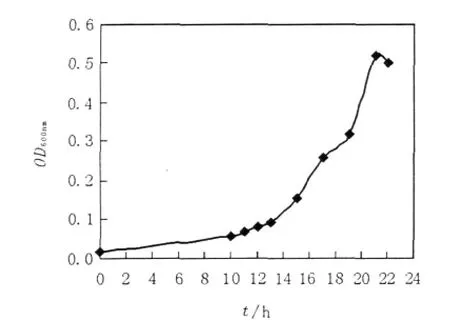
圖6 乙酸鹽為唯一碳源條件下M.smegma菌生長曲線Fig.6The growth curve of M.smegmatis when acetate as the sole carbon resource
同樣,考察DMSO濃度對M.smegma生長的影響,發現當DMSO終濃度(體積比)為1%時對菌體無明顯的抑制作用,但隨DMSO濃度的增加,其抑制作用逐漸增加,特別是當DMSO終濃度達到4%時,菌體生長抑制率約為20%,故確定篩選體系中DMSO終濃度不超過1%。最終確定乙醛酸代謝途徑抑制劑的細胞水平篩選體系:在198 μL M.smegma 15 h培養物中加入2 μL待篩選樣品溶液。
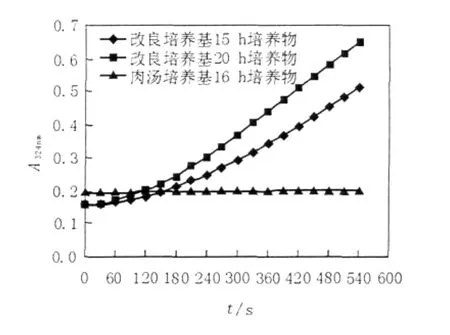
圖7 乙酸鹽為唯一碳源條件下M.smegma內ICL活性Fig.7The intracellular activity of ICL of M.smegmatis
2.7 乙醛酸途徑抑制劑細胞水平篩選步驟
于96孔板中,每孔加入2 μL待篩選樣品溶液,198 μL M.smegma在改良Dubos培養基中15 h培養物,在37℃溫箱內震蕩3 min后于600 nm測定OD值,計為OD1,繼續在37℃溫箱內震蕩培養5 h,于600 nm測定OD值,計為OD2。同時以2 μL DMSO代替待篩選樣品作為陰性對照,檢測其培養過程中吸光度變化值(OD2-OD1)記做Δ對照(OD2-OD1)。
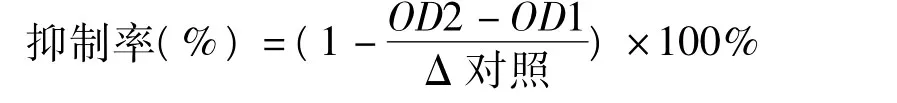
2.8 樣品的篩選
取微生物發酵次級代謝產物樣品1060個,利用ICL酶水平抑制劑篩選模型進行初篩,獲得10個陽性樣品,再利用乙醛酸途徑抑制劑細胞水平篩選模型進行復篩,復篩的陽性率(抑制率達到50%以上)可達到90%(表1),說明2種模型的相關性較好。
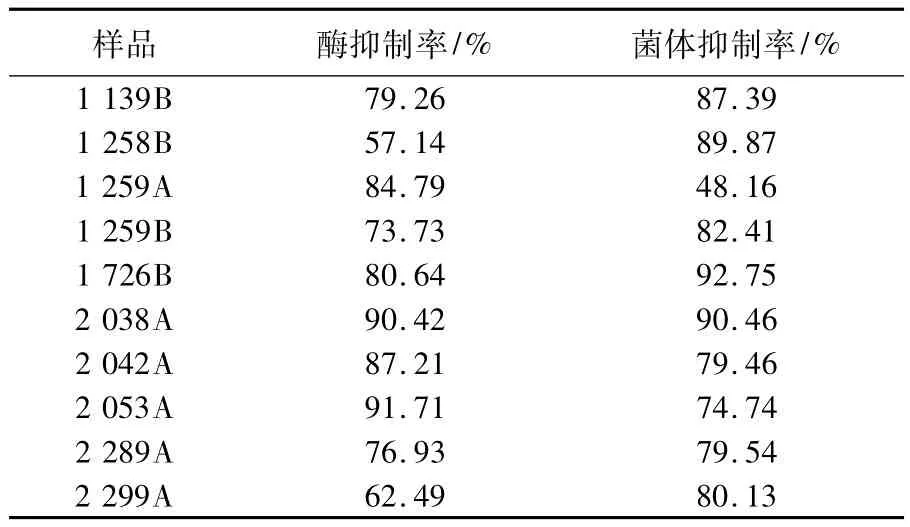
表1 不同樣品對ICL和乙醛酸代謝途徑的抑制率Table 1The inhibition ratio to ICL and glyoxalic acid pathway of differentive samples
3 討論
異檸檬酸裂解酶(ICL)是乙醛酸循環的關鍵酶,對M.tuberculosis在巨噬細胞內的持留狀態起關鍵作用[3]。以異檸檬酸裂解酶作為靶點篩選得到的抗結核藥物不僅可以有效殺滅處于持留狀態的M.tuberculosis,縮短結核病治療療程,同時由于哺乳動物體內不存在乙醛酸循環,因此以ICL為靶點開發的新藥毒副作用較小。實驗中針對ICL為靶點,先在酶(分子)水平后又在菌體(細胞)水平建立篩選模型,不僅是對樣品活性的驗證,還使篩選模型更接近于持留狀態的M.tuberculosis,篩選結果更具有真實性、可靠性。
為了使ICL抑制劑篩選模型有較高的靈敏性,本實驗中采用最適的酶促條件進行篩選反應,使反應體系敏感度高。在滿足篩選要求的前提下,使用最少的酶量,以增加低濃度抑制劑的檢出率。由于樣品為微生物發酵液乙酸乙酯萃取物,成分復雜,在水相中溶解效果較差,會影響ICL抑制劑篩選模型的穩定性和可靠性。通過對多種溶劑的考察和對難溶樣品的處理,模型的穩定性和可靠性明顯增強。
Kerstin H?ner Zu Bentrup等研究表明,使用改良的Dubos培養基可使M.tuberculosis利用乙酸鹽作為唯一碳源,ICL表達水平上調,為在細胞水平上ICL抑制劑的篩選奠定基礎。由于M.tuberculosis具有高致病性、高傳染性,實驗條件苛刻且生長速度緩慢等缺點,不利于大規模高通量篩選的實現,而同為分枝桿菌屬的M.smegma為非致病性菌株,且生長周期較短,僅為3~4 h[7];另一方面,Vinita Chaturvedi等[8]評價了以M.smegma作為M.tuberculosis替代菌株的可行性,認為可以使用M.smegma進行MDR-TB藥物的篩選。2001年開始的M.smegma基因組測序,發現M.smegma基因組中也存在異檸檬酸裂解酶基因,比較M.tuberculosis ICL序列和M.smegma ICL序列,可發現二者有很高的相似性。本實驗表明,M.smegma在改良的Dubos培養基上也可使用乙酸鹽為唯一碳源生長,因此本實驗中使用M.smegma作為替代菌株,進行乙醛酸代謝途徑抑制劑的篩選。
Sharma V等[9]發現ICL抑制劑3-硝基丙酸鹽在以乙酸鹽為唯一碳源的培養基上對野生型M.smegma以及基因重組的M.smegma(使用M.tuberculosis ICL基因替換野生型M.smegma基因組中的ICL基因)均有較強的抑制作用,而這一現象并未在以葡萄糖為唯一碳源的培養條件下觀測到,證明了乙醛酸途徑抑制劑細胞水平篩選模型的可行性,并在一定程度說明了酶水平篩選與菌體水平篩選的相關性。
[1]Cole ST,Brosch R,Parkhill J,et al.Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence[J].Nature,1998,393(6685):537-544.
[2]Kerstin H?ner Zu Bentrup,Andras Miczak,Dana L.Swenson,et al.Characterization of Activity and Expression of Isocitrate Lyase in Mycobacterium avium and Mycobacterium tuberculosis[J].Bacteriol,1999,181(23):7161-7167.
[3]McKinney J D,Bentrup K H Z,Munoz-Elias E J,et al.Persistence of Mycobacterium tuberculosias in macropha ge and mice requires the glyoxylate shunt enzyme isocitrate lyase[J].Nature,2000,406(6797):735-738.
[4]王旸,肖春玲.以異檸檬酸裂解酶為靶點篩選抗持留結核分枝桿菌藥物[J].中國抗生素雜志,2007,32(7):391-395.
[5]賴煦卉,王洪海,張雪蓮.異檸檬酸裂解酶—新型抗結核藥物靶標[J].中國感染控制雜志,2008,7(5):359-362.
[6]Ranaldi F,Vanni P,Giachetti E.Enzyme catalysis in microgravity:steady-state kinetic analysis of the isocitrate lyase reaction[J].Biophys Chem,2003,103(2):169-77.
[7]Reyrat JM,Kahn D Mycobacterium smegmatis:an absurd model for tuberculosis[J].Trends Microbiol,2001,9(10):472-473.
[8]Chaturvedi V,Dwivedi N,Tripathi RP,et al.Evaluation of Mycobacterium smegmatis as a possible surrogate screen for selecting molecules active against multi-drug resistant Mycobacterium tuberculosis[J].J Gen Appl Microbiol,2007,53(6):333-337.
[9]Sharma V,Sharma S,Hoener zu Bentrup K,et al.Structure of isocitrate lyase,a persistence factor of Mycobacterium tuberculosis[J].Nat Struct Biol,2000,7(8):663-668.
Establishment of New Screening Models for Anti-Mycobacterium tuberculosis Medicines
ZHANG Lei-lei1,YOU Peng-fei1,2,YAO Guang-xin1,HOU Jue1,ZHANG Yi-xuan1,2
(1.Teach.&Res.Div.of Bio-Pharm.,2.Biol.&Exp Teach.Ctr.,Shenyang Pharm.Uni.,Shenyang 110016)
Mycobacterium tuberculosis strain H37Rv genomic DNA was adopted as a template to establish a screening model targeting to isocitrate lyase(ICL)and glyoxylate pathway for anti-TB drugs at enzymatic and cellular levels.ICL gene was amplified by polymerase chain reaction(PCR)and constructed an expression vector and highly expressed in E.coli BL21(DE3).The recombinant ICL was purified with Ni2+-NTA resin affinity chromatography and detected for its activity.The ICL enzymatic reaction conditions were optimized,and observed the effects of solvents on the enzymatic activity of the samples of going to screen to establish a screening model at the level of inhibition enzyme; observed and optimized the growth conditions of M.smegmatis(Ms)in medium with acetate as a sole carbon source,to establish a screening model at cellular level based on inhibitor of glyoxylate pathway of Ms.The results showed that using the two screening models above-mentioned 1060 kinds of microbial metabolite samples in the lab possibly possessing antagonistic activity were screened initially and rescreened,and both the screened results had fairly good positive reciprocity.
Mycobacterium tuberculosis;isocitrate lyase;Mycobacterium smegmatis;glyoxylate pathway;screening model
Q939-92
A
1005-7021(2011)01-0057-06
國家重大新藥創制專項(2009ZX09301-012)資助
張磊磊男,碩士。研究方向為微生物活性代謝產物的篩選。E-mail:zhll1986211@163.com
*通訊作者。Tel:024-23984102,E-mail:zhangyxzsh@163.com
2010-12-25;
2011-01-21
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
光學精密工程(2016年6期)2016-11-07 09:07:19
學苑創造·A版(2015年11期)2016-01-14 09:03:27
核科學與工程(2015年4期)2015-09-26 11:59:03
中國火炬(2010年8期)2010-07-25 11:34:30
