幽門螺桿菌對甲硝唑的耐藥性分析*
2011-08-21 06:51:52孫朝琴羅昭遜夏曙華
中國人獸共患病學報 2011年12期
張 姝,莫 非,孫朝琴,張 然,王 瑩,李 寅,羅昭遜,夏曙華
幽門螺桿菌是淺表性胃炎、消化性潰瘍的致病因子,與胃癌、胃粘膜相關性淋巴組織淋巴瘤的發生密切相關[1]。目前治療和控制Hp感染的主要手段是抗生素治療,在治療Hp感染的抗生素中以甲硝唑應用最廣,隨著甲硝唑的廣泛應用,其耐藥性問題日益突出,已成為治療失敗的主要原因之一[2]。Hp對甲硝唑耐藥機制尚未完全明了,目前主要認為對氧不敏感的NADPH硝基還原酶(Rdx A)失活是導致甲硝唑耐藥的關鍵,該酶的編碼基因為rdx A基因[3]。有文獻報道參與Hp對甲硝唑耐藥的硝基還原酶還包括:3種鐵氧還蛋白類似物(Fdx B)、NADPH 黃素氧還酶(Frx A)等[4-5]。但在 Hp耐甲硝唑機制的研究中,對fdx B和frx A的報道甚少,本文擬采用PCR擴增rdx A、fdxB和frx A基因,并對這3個基因測序分析,探討Hp對甲硝唑耐藥與rdx A、frx A和fdx B點突變的關系。
1 材料與方法
1.1 菌株來源 采自貴陽醫學院附屬醫院,60例患者因胃部不適行胃鏡檢查,標本取自其胃竇部或胃體部粘膜,并且C14尿素呼氣試驗為陽性的情況下,進行Hp的分離培養所得。國際標準測序株Hp26695株購自ATCC。
1.2 主要試劑 E-Test試驗條、微需氧袋:法國梅里埃公司;哥倫比亞血瓊脂粉:OXOID;DNA提取試劑盒:北京天恩澤基因科技有限公司。
1.3 Hp的分離培養及鑒定 取胃竇部內鏡活檢組織,接種于幽門螺桿菌選擇培養基平板,微需氧37℃培養3~7 d,進行Hp鑒定及培養。
1.4 藥敏試驗 采用E-Test法檢測Hp對甲硝唑的最低抑菌濃度(MIC)。用 Hp標準測序株Hp26695作每批試驗的質控標準。以MIC值≥8 μg/m L判定為甲硝唑耐藥,MIC值≥32μg/m L判定為高水平甲硝唑耐藥[6]。
1.5 Hp基因組DNA提取 提取DNA按照試劑盒說明書操作。用紫外分光光度計對提取的DNA純度進行測定,計算 OD260nm/OD280nm比值。模板DNA于1.0%瓊脂糖凝膠孔中電泳。
1.6 PCR 擴增 根據文獻[7]設計rdx A、fdx B和frx A基因的特異性引物,見表1。引物由上海生物工程有限公司合成。PCR反應體系為反應體系為50μL,其中包括:dd H2O 19μL,上游引2μL,下游引物2μL,模板DNA 2μL,Primix Teq 25μL。循環條件:預變形94℃,5 min;變性94℃,30 s;退火53.7℃(其中fdx B退火59℃~60℃,frx A退火59℃~60℃);延伸72℃2 min;最終延伸72℃,10 min。每次擴增時均設置陽性對照、陰性對照及空白對照。最后瓊脂糖凝膠電泳檢測PCR產物。
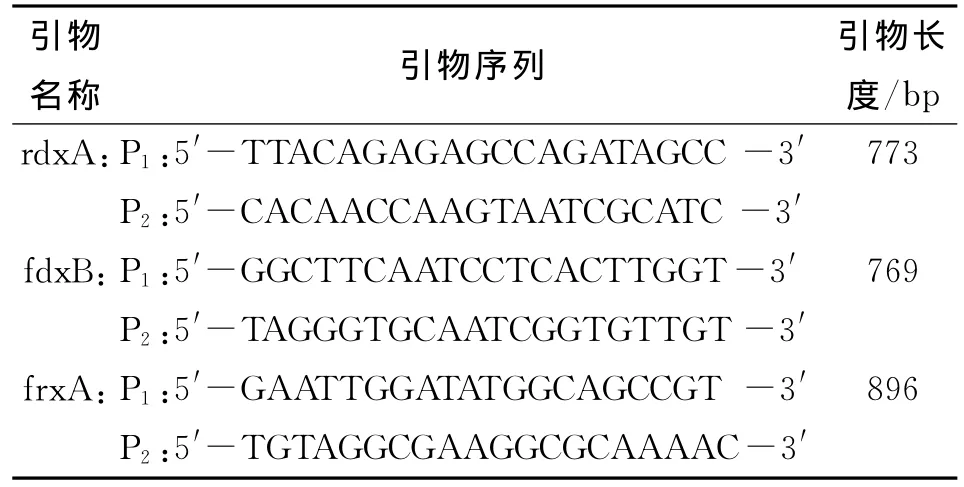
表1 rdx A、fdxB、frx A引物序列Tab.1 Primers of rdxA,fdxB and frx A
1.7 PCR產物測序及序列分析 PCR產物送北京三博遠志生物技術有限責任公司測序。測序結果經美國國立生物技術信息中心(NCBI)中的BLAST軟件分析。
2 結 果
2.1 Hp的培養及藥敏結果 60例病人胃粘膜標本分離出11株Hp,經革蘭氏染色鏡檢呈陰性,菌體細長彎曲呈螺形,S型或海鷗型。過氧化氫酶實驗、氧化酶實驗及尿素酶實驗,均為陽性。采用ETest法測定11株臨床株對甲硝唑的MIC均≥256 μg/m L,為高水平耐藥株。
2.2 PCR擴增結果 11株臨床株和國際標準測序株Hp26695的DNA純度經紫外分光光度計測定,260 nm/280 nm比值均在1.8~2.0之間。PCR擴增出11株臨床株和1株標準株rdx A、frx A和fdxB基因,分別在750 bp、870 bp、830 bp位置出現特異條帶,見圖1。
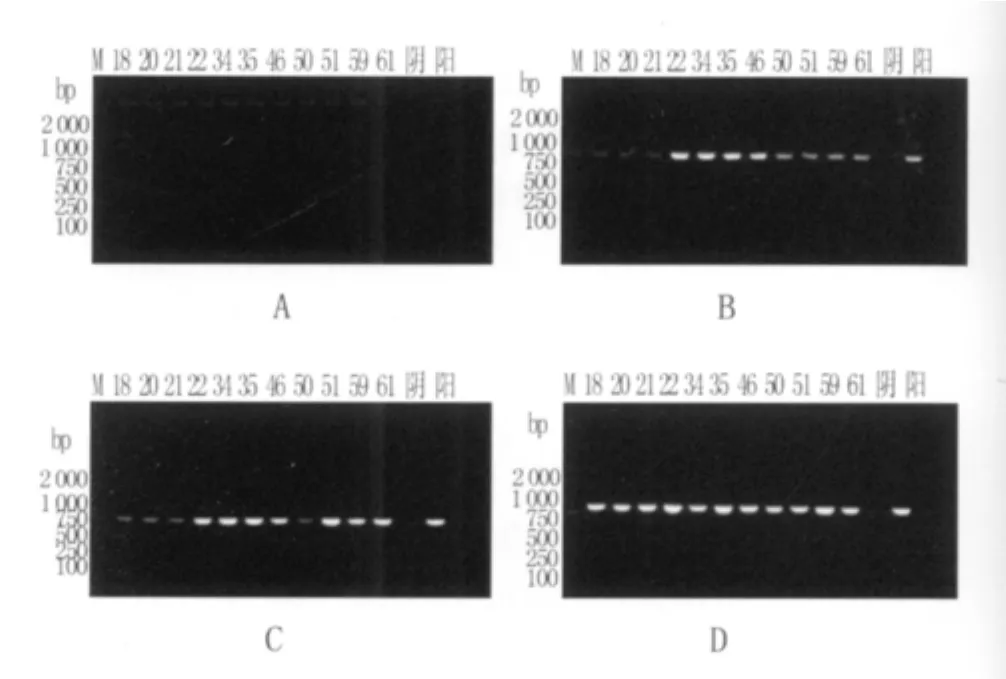
圖1 模板DNA及PCR擴增產物的瓊脂糖凝膠電泳圖注:A圖為模板DNA;B圖為rdx A;C圖為fdxB;D圖為frx A;M:DNA marker;陰:陰性對照;陽:標準株Hp26695;其余為臨床株。Fig.1 Agarose gel electrophoresis maps of DNA templates and PCR amplification productsNote:Picture A for DNA templates;B for rdx A;C for fdxB;D for frx A;M for marker,positive for the standard strain Hp26695,the rest for clinical strains
2.3 產物測序結果 11株臨床株與NCBI已登錄的國際標準測序株Hp26695相應基因組序列比對,發現rdx A、fdx B、frx A基因存在堿基替換、插入和缺失3種類型的突變,這些基因突變呈現出一定的規律性,即在某些位點存在共同突變(共同突變位點見表1),除此,還存在隨機位點的散在突變。耐藥菌與敏感菌核苷酸同源性:rdx A和fdxB為95%~98%,frx A低于94%。
3 討 論
硝基咪唑類藥物甲硝唑(MTZ)是一種具有抑菌活性的藥物前體,經Hp的細胞膜被動擴散進入胞漿,在胞漿中的硝基還原酶的作用下,能獲得低于-430 m V氧化還原電位,使甲硝唑的硝基還原成羥胺衍生物,還原產物與DNA作用引起鏈斷裂,導致細菌死亡,這是甲硝唑的抗Hp機制[3,8]。因其殺菌活性不受胃內低p H的影響,在胃腔內濃度高,具有較強的抗Hp活性,因此常用于Hp感染的一線治療[9],是三聯方案中最廣泛應用的抗菌素之一。隨著抗生素的廣泛使用,Hp對甲硝唑的耐藥率呈逐年上升趨勢。本實驗從60例病人胃粘膜標本分離出11株Hp菌株,采用E-Test法測定11株臨床株對甲硝唑的MIC均≥256μg/m L,均為高水平耐藥株。
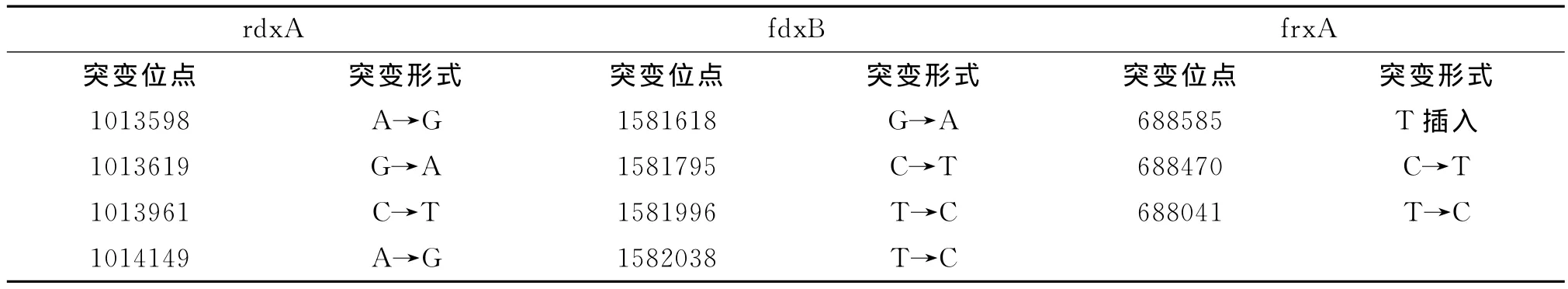
表2 11株耐藥菌rdx A、fdxB和frxA基因的相同突變位點和突變形式Tab.2 The same mutations sites and forms of rdxA,fdxB and frx A in 11 resistant strains
Kwon等研究表明Hp對甲硝唑耐藥與編碼參與甲硝唑氧化還原反應的酶的基因(rdx A、frx A、fdx B)突變有關[10]。這些基因的堿基插入、缺失、轉化等導致下游氨基酸序列全部或部分發生改變,導致其翻譯的蛋白質改變,引起硝基還原酶活性降低或失活,不能還原甲硝唑而引起耐藥。但對氧不敏感的NADPH硝基還原酶(Rdx A)最為關鍵,國外有研究表明,耐甲硝唑的Hp中的Rdx A蛋白缺失或生成減少[11-12]。本研究對11株耐藥菌和1株敏感菌進行rdx A擴增并測序,發現11株耐藥菌均存在插入、缺失和堿基轉換3種突變類型,以堿基轉換為突變的主要類型,未發現大片段序列插入或缺失,只出現單個堿基插入或缺失,即均表現為點突變。已有報道認為耐甲硝唑Hp菌株的rdx A基因突變大多為隨機性的,未能肯定某一位點突變與耐藥直接相關[13]。本試驗也發現rdx A基因變異位點大多不固定,但11株耐藥菌存在4個固定的突變位點,即均發生了1013598nt A→G,1013619nt G→A,1013961nt C→T,1014149nt A→G,這些位點的改變均未見文獻報道,我們推測這些位點的突變與甲硝唑的耐藥可能有密切的關系。
Hp的frx A基因和fdx B基因的變異在Hp對甲硝唑中-高度耐藥中發揮重要作用[4]。有研究證實單一fdx B基因突變導致Hp對甲硝唑的低水平的耐藥甚至不耐藥,rdx A或frx A基因突變會產生中等程度的耐藥,但當rdx A與fdx B或frx A聯合突變則會產生高水平耐藥[10,14]。本文對11株高水平耐藥菌的frx A基因和fdx B基因進行擴增和測序分析,發現耐藥株和標準株均攜帶有frx A和fdx B基因,且發生不同程度的變異,多表現為隨機位點的插入、缺失、轉化及顛換,未發現大片基因的插入,但是同時也呈現出一些規律性的變化,如:frx A基因測序結果在以下3個位點出現了共同突變:688585nt T插入,688470nt C→T,688041nt T→C,同源性為95%~98%;fdx B的基因測序結果表明,11株高水平的耐藥株均發生變異,有以下共同突變位點,即 1581618nt處 G→A,1581795ntC→T,1581996nt T→C,1582038nt T→C,同源性低于94%。我們推測這些高頻的突變位點可能在該基因所編碼的蛋白產物失活中起重要作用。從本研究的實驗結果來看,11株高水平耐甲硝唑的Hp菌同時發生了rdx A、frx A和fdx B基因突變,這在一定程度上證實了frx A和fdx B基因突變在rdx A基因突變所誘導的甲硝唑耐藥中起協同作用。frx A基因和fdx B在甲硝唑耐藥中是否單獨起作用還需要增加甲硝唑敏感株,需要更多的實驗數據去做進一步的研究。
研究Hp對甲硝唑的耐藥機制可揭示藥物作用細菌后信號轉導機制,為個體化用藥以及選擇合適的抗生素用于臨床治療奠定基礎。本文從基因組角度探討Hp對甲硝唑耐藥的分子機制,提示了Hp對甲硝唑耐藥性不僅可能與rdx A點突變有關,同時亦可能與frx A和fdxB基因突變相關。為進一步研究和分析Hp耐藥機制提供了有價值的資料。
[1]Cover TL,Blaser MJ.Helicobacterpyloriin health and disease J[J].Gastroenterology,2009,136(6):1863-1873.
[2]Weel JEL,van der Hulst R,Gerrists Y,et al.Heterogeneity in metronidazole amongHelicobacacterpyloriisolatesμLcer disesse[J].J Clin Microbiol,1996,34(9):2158-2162.
[3]張燕捷,吳叔明.幽門螺桿菌耐藥性的研究現狀[J].上海第二醫科大學學報,2005,25:93-96.
[4]Goodwin A,Kersulyte D,Sissnon G,et al.Metronidazole resistance inHelicobacterpyloriis due to null mutations in a gene(rdx A)that encodes an oxgen-insentive NADPH nitroreductase[J].J Mol Microbiol,1998,28(2):383-393.
[5]Solca NM,Bernasconi MV,Piffaretti JC.Mechanism of metronidazole resistance inHelicobacterpylori:comparison of the rdx A gene sequences in 30 strains[J].Antimicrob Agents Chemother,2000,44:2207-2210.
[6]Jeong JY,Mukhopadhyay AK,Dailidiene D,et al.Sequential inactivation of rdx A(HP0954)and frx A (HP0642)nitroreductase genes causesmoderate and high-level metronidazole resistance inHelicobacterpylori[J].J Bacteriol,2000,182(18):5082-5090.
[7]李耿,田擁軍,葉嗣穎,等.幽門螺桿菌對甲硝唑耐藥性及多重相關基因DNA和氨基酸序列分析[J].華中科技大學學報,2005,34(2):138-140.
[8]Cederbrant G,Kahlmeter G,Ljungh A.Propsed mechanism for metronidazole resistrance inHelicobacterpylori[J].J Antimicrob Chemother,1992,29(2):115-120.
[9]單蘊,徐樹長,葉元康.幽門螺桿菌抗生素耐藥率分析[J].同濟大學學報:醫學版,2004,25(1):31-33.
[10]Kwon DH,EI-Zaatari FA,Kato M,et al.Analysis of rdx A and involvement of additional genes encoding NAD(P)H flavin oxidoreductase(Frx A)and ferredoxin-like protein(Fdx B)in metronidazole resistance ofHelicobacterpylori[J].Antimicrob A-gents Chemother,2000,44(8):2133-2142.
[11]Latham SR,Labigne A,Jenks PJ.Production of the Rdx A protein in metronidazole-susceptible and resistant isolates ofHelicobacter pyloricultured from treated mice[J].J Antimicrob Chemother 2002,49(40):675-678.
[12]Latham SR,Owen RJ,Wlviss NC,et al.Differentiation of metronidazole-sensitive and resistant clinical isolates ofHelicobacter pyloriby immunoblotting with antisera to the Rdx A protcin[J].Clin Microbiol,200l,39:3025-3055.
[13]Solca NM,Bemasconi MV,Piffaretti JC.Mechanism of metronidazole resistance inHelicobacterpylori:comparison of rdx A gene sequences in 30 strains[J].Antimicrob Agents Chemother,2000,44(8):2207-2210.
[14]Marais A,Bilardi C,Cantet F,et al.Characterization of the genes rdx A and frx A involved in metronidazole resistance inHelicobacterpylori[J].Res Microbiol,2003,154(2):137-144.
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
中國生殖健康(2020年2期)2021-01-18 02:51:26
科學大眾(2020年12期)2020-08-13 03:22:22
小學生導刊(2018年13期)2018-06-29 03:49:00
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
