肌肉來源干細胞表面抗原-1陽性與陰性細胞體外培養成肌特性的比較
2011-07-17 07:30:56車曉霞趙彤朱玲玲郭杰
華西口腔醫學雜志 2011年4期
車曉霞 趙彤 朱玲玲 郭杰
(1.首都醫科大學附屬北京口腔醫院 正畸科,北京 100050;2.軍事醫學科學院基礎醫學研究所 腦保護與可塑性研究室,北京 100850;3.山東大學口腔醫院 正畸科;山東省口腔生物醫學重點實驗室,濟南 250012)
干細胞表面抗原-1(stem cell antigen-1,Sca-1)作為一種細胞表面標記常用于富集干細胞和前體細胞,最早發現于小鼠激活的淋巴細胞[1],是一種由鼠Ly6基因家族編碼,以糖基磷脂酰肌醇錨定的細胞表面蛋白,相對分子質量為18 000,定位于漿膜的脂質載體[2]。
骨骼肌受到損傷后,在骨骼肌局部存留的干細胞和肌肉形成前體細胞激活、增殖、分化并融合形成多核肌細胞,修復或替代受損的組織[3-4]。這對于維持骨骼肌在運動和老化過程中的動態平衡非常重要。實驗證實表達Sca-1的細胞在體內和體外都可以分化為骨骼肌,也可以生成造血細胞和間充質細胞[5-6]。在細胞移植治療中,Sca-1+細胞存活率高[7],靶基因治療效果好,但目前對這種細胞表面蛋白的功能尚不明了。
成肌調節因子(muscle regulatory factor,MRF)在肌肉細胞種系決定和分化中起重要作用。其中MyoD和成肌素(Myogenin)都屬于MRF,MyoD作為原發的轉錄因子,在成肌種系決定中起作用[8],見于激活的衛星細胞;Myogenin作為繼發的轉錄因子,在肌肉終末分化時表達[9]。本研究對比觀察了Sca-1+與Sca-1-細胞在體外培養過程中的生長特性及MRF表達的特點,希望獲得實驗線索以說明Sca-1在細胞增殖分化過程中所發揮的功能作用。
1 材料和方法
1.1 小鼠原代成肌細胞的分離和培養
將4周齡C57BL/6小鼠脫頸處死,75%乙醇消毒小鼠后肢皮膚,剪開皮膚,剔去脂肪和筋膜,剪取后肢肌肉組織,立即放入預冷的DMEM中漂洗3次。將肌肉小塊轉移至無菌的小燒杯中,用眼科剪充分剪碎成漿狀,加入組織混合酶(2.4 U·mL-1分散酶、0.2%Ⅱ型膠原酶、2.5mmol·L-1CaCl2),37℃消化45min,每間隔15min用吸管反復吹吸若干次。用含20%胎牛血清(fetal bovine serum,FBS)的生長液終止消化,200目細胞篩選過濾,1 400 r·min-1離心10min,棄去上清,按照密度為每毫升1×106個細胞懸于200μL的PBS中,等待流式細胞儀檢測和分選。
1.2 流式細胞儀檢測原代成肌細胞中Sca-1的表達
將從小鼠骨骼肌中獲得的新鮮成肌細胞以密度為每毫升1×106個懸于200μL的PBS中,加入藻紅蛋白標記的Sca-1單克隆抗體(Santa Cruz Biotechnologies公司,美國),37℃孵育30min,用含體積分數0.1%牛血清蛋白(bovine serum albumin,BSA)的PBS洗滌、重懸細胞,移入專用測試管,應用流式細胞儀檢測Sca-1的表達,采用聯機專用軟件Cell Quest進行分析。此外按照流式細胞儀儀器手冊設置各項參數,安裝分選器,將上述細胞進行分選,分別收集Sca-1+與Sca-1-細胞。
1.3 生長曲線的測定
生長曲線是判定細胞活力的重要指標,本實驗采取CCK-8試劑盒測定光密度值(A值),間接測量活細胞數量。將流式細胞儀分選獲得的Sca-1+與Sca-1-細胞分別以密度為每孔3×104個細胞接種于96孔板,每組20孔。培養1、3、5 d后向各孔中加入10μL的CCK-8溶液,37℃孵育4 h,直接測定450 nm處的A值,繪制曲線。
1.4 Western blot檢測成肌蛋白的表達
為了對比小鼠骨骼肌來源Sca-1+與Sca-1-細胞在體外培養過程中從增殖到分化的變化,采用Western blot檢測Sca-1、MyoD、Myogenin的蛋白表達。將Sca-1+與Sca-1-細胞分別以密度為每毫升5×104個細胞接種到涂布過鼠尾膠原的60mm培養皿中,培養液為高糖DMEM(含有20%FBS、1%青鏈霉素、2.5 ng·mL-1堿性成纖維細胞生長因子),細胞培養4 d后用細胞刮刮下細胞,PBS清洗,離心,加入細胞蛋白裂解液,冰上裂解30min,提取蛋白,-80℃保存備用。
常規制備10%SDS-PAGE分離膠,混勻后,將其灌入預先制備好的制膠板內(Bio-Rad公司的Mini-Cell制膠系統),用水覆蓋,室溫凝固30min。制備5%SDS-PAGE濃縮膠,混勻后,灌膠并置室溫凝固30min。電泳首先以8 V·cm-1進行,待上樣緩沖液進入分離膠后調整為15V·cm-1,電泳時間約2 h;隨后以60 V的恒壓進行硝酸纖維素膜電轉,電轉時間視被轉移蛋白的大小而定。電轉結束后,將膜置于TBS中漂洗,5%脫脂奶粉(北京普利萊公司)封閉,經TBS充分振蕩清洗后,分別滴加一、二抗(辣根過氧化物酶標記的相應抗體),DAB顯色,觀察結果。
2 結果
2.1 Sca-1+與Sca-1-細胞體外培養增殖能力的測定結果
流式細胞儀檢測原代細胞中Sca-1的陽性表達率為64.86%,分選后分別進行Sca-1+與Sca-1-細胞的體外培養,經CCK-8試劑盒測定2種細胞的生長曲線見圖1。在培養過程中,前3 d Sca-1+與Sca-1-細胞增殖速率沒有明顯差別,第3天后Sca-1-細胞開始加速增殖。

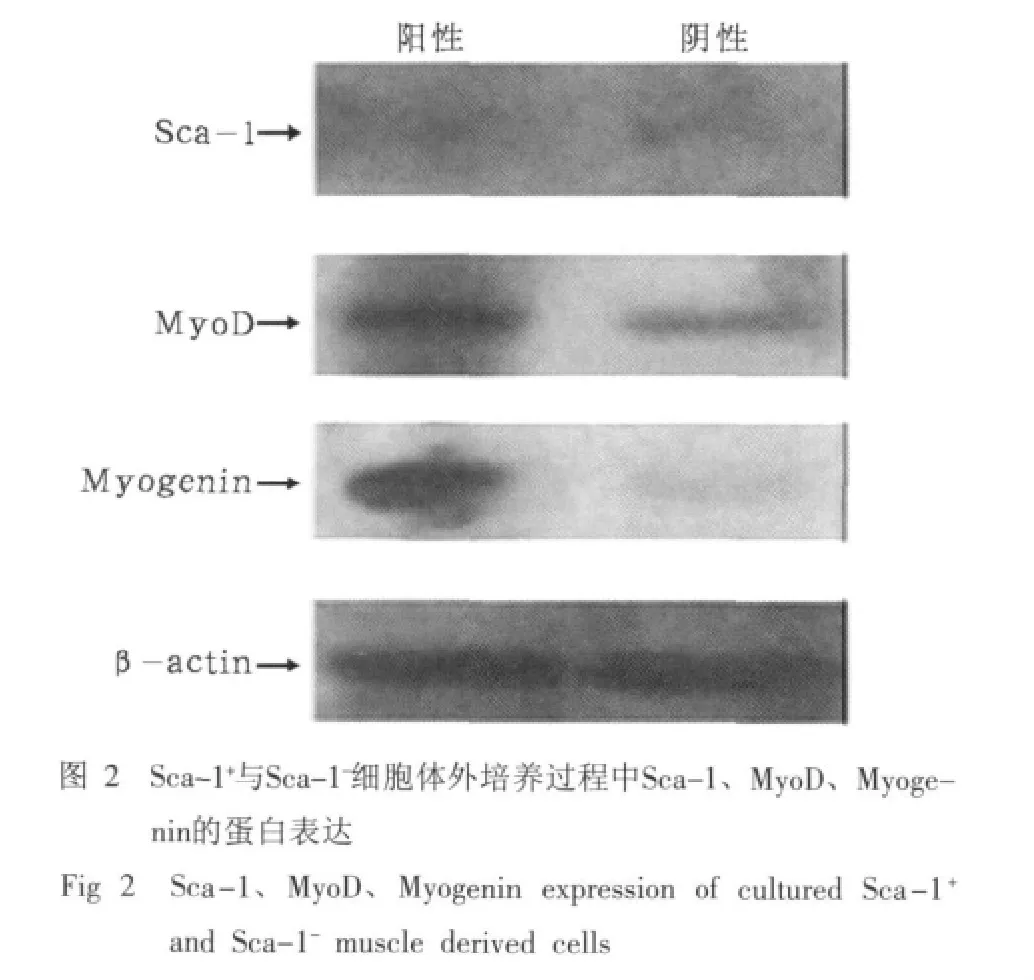
2.2 Sca-1+與Sca-1-細胞體外培養過程中Sca-1、Myo-D、Myogenin的蛋白表達
Sca-1+與Sca-1-細胞體外培養過程中Sca-1、Myo-D、Myogenin的蛋白表達結果見圖2。由圖2可見,Sca-1+與Sca-1-細胞在體外培養過程中Sca-1表達均不明顯,Sca-1+細胞比Sca-1-細胞MyoD、Myogenin表達強,說明Sca-1+比Sca-1-細胞在體外培養過程中具有更好的成肌特性。
3 討論
從不同的組織中都可以分離出具有干細胞特征的Sca-1+細胞[10]。在體外培養骨骼肌來源成肌細胞的過程中發現,Sca-1+與Sca-1-細胞在體外培養5 d的過程中Sca-1均未見明顯表達。Jankowski等[11]研究也顯示細胞表面蛋白的表達在體外培養過程中會發生變化。隨著培養時間的延長,Sca-1表達下降。以Sca-1表達為標記純化出來的成肌細胞體外培養5 d,Sca-1表達消失。這些結果說明,Sca-1的出現是細胞生長發育過程中一個時段的表達特征,以此作為標記進行干細胞篩選可能會產生一定的局限性。
盡管對于Sca-1在細胞生命活動的作用尚不了解,但是很多實驗結果卻支持Sca-1的細胞信號功能[12-13]。Sca-1基因沉默小鼠和骨骼肌成肌細胞的實驗顯示Sca-1在成肌細胞增殖、分化、融合中發揮作用。對于體外培養的C2C12成肌細胞,給予Sca-1抗體阻斷,其功能會促進細胞增殖和抑制成肌細胞融合。從體內實驗角度來看,分別在Sca-1+和Sca-1-鼠骨骼肌中形成壞死性損傷,Sca-1-成肌細胞出現持續和加速的細胞分裂,表現為高度的增殖反應[14]。本實驗體外培養增殖能力的檢測結果也說明Sca-1-比Sca-1+細胞具有更強的增殖能力。
Sca-1對于成肌細胞增殖分化的作用可能與對細胞進出細胞周期的影響有關。通過基因芯片實驗,學者們發現在成肌細胞早期分化進行過程中,細胞離開細胞循環時,包括DDAH2和Sca-1等幾個基因表達特異性上調[12];當細胞開始分化時Sca-1表達出現暫時性升高[14]。有學者認為,成肌調節因子的表達與細胞分裂相排斥[13],MyoD的功能是通過誘導細胞周期蛋白依賴性激酶抑制劑p21[15-16],同時伴隨其他Cip1/Kip1家族成員p57和p27,廣泛抑制細胞周期蛋白依賴性激酶的活性,從而使細胞停止分裂[17]。Myogenin則在肌肉終末分化時表達[9]。本實驗結果顯示:Sca-1+比Sca-1-細胞表達更多的MyoD和Myogenin。從以往的研究結果可以推測得出解釋:雖然從骨骼肌中分離出來的成肌細胞表達Sca-1,但其中一部分可能是已經走出細胞周期,開始了早期分化細胞,因而直接表達成肌分化蛋白;而Sca-1-成肌細胞表現出的高度增殖反應導致成肌細胞分化延遲。這個結果進一步提示,如果以Sca-1為標記分離出來的細胞可能是還沒有進入細胞循環的干細胞,或者是已經進入早期分化階段的成肌細胞。
[1]Yutoku M,Grossberg AL,Pressman D.A cell surface antigenic determinant present on mouse plasmacytes and only about half of mouse thymocytes[J].J Immunol,1974,112(5):1774-1781.
[2]Stefanová I,HorejsíV,Ansotegui IJ,et al.GPI-anchored cellsurface molecules complexed to protein tyrosine kinases[J].Science,1991,254(5034):1016-1019.
[3]ChargéSB,Rudnicki MA.Cellular and molecular regulation of muscle regeneration[J].Physiol Rev,2004,84(1):209-238.
[4]Dhawan J,Rando TA.Stem cells in postnatal myogenesis:Molecular mechanisms of satellite cell quiescence,activation and replenishment[J].Trends Cell Biol,2005,15(12):666-673.
[5]Lee JY,Qu-Petersen Z,Cao B,et al.Clonal isolation of musclederived cells capable of enhancing muscle regeneration and bone healing[J].J Cell Biol,2000,150(5):1085-1100.
[6]Royer CL,Howell JC,Morrison PR,et al.Muscle-derived CD45-SCA-1+c-kit-progenitor cells give rise to skeletal muscle myotubes in vitro[J].In Vitro Cell Dev Biol Anim,2002,38(9):512-517.
[7]Gussoni E,Soneoka Y,Strickland CD,et al.Dystrophin expression in the mdx mouse restored by stem cell transplantation[J].Nature,1999,401(6751):390-394.
[8]Zhang W,Behringer RR,Olson EN.Inactivation of the myogenic bHLH gene MRF4 results in up-regulation of myogenin and rib anomalies[J].Genes Dev,1995,9(11):1388-1399.
[9]Cornelison DD,Wold BJ.Single-cell analysis of regulatory gene expression in quiescent and activated mouse skeletalmuscle satellite cells[J].Dev Biol,1997,191(2):270-283.
[10]Holmes C,Stanford WL.Concise review:Stem cell antigen-1,Expression,function,and enigma[J].Stem Cells,2007,25(6):1339-1347.
[11]Jankowski RJ,Haluszczak C,Trucco M,et al.Flow cytometric characterization ofmyogenic cell populations obtained via the preplate technique:Potential for rapid isolation of muscle-derived stem cells[J].Hum Gene Ther,2001,12(6):619-628.
[12]Shen X,Collier JM,Hlaing M,et al.Genome-wide examination ofmyoblast cell cycle withdrawal during differentiation[J].Dev Dyn,2003,226(1):128-138.
[13]Smith TH,Block NE,Rhodes SJ,et al.A unique pattern of expression of the four muscle regulatory factor proteins distinguishes somitic from embryonic,fetal and newborn mouse myogenic cells[J].Development,1993,117(3):1125-1133.
[14]Epting CL,López JE,Pedersen A,et al.Stem cell antigen-1 regulates the tempo of muscle repair through effects on proliferation of alpha7 integrin-expressing myoblasts[J].Exp Cell Res,2008,314(5):1125-1135.
[15]Guo K,Wang J,Andrés V,et al.MyoD-induced expression of p21 inhibits cyclin-dependent kinase activity upon myocyte terminal differentiation[J].Mol Cell Biol,1995,15(7):3823-3829.
[16]Halevy O,Novitch BG,Spicer DB,et al.Correlation of terminal cell cycle arrest of skeletalmuscle with induction of p21 by MyoD[J].Science,1995,267(5200):1018-1021.
[17]Sherr CJ,Roberts JM.CDK inhibitors:Positive and negative regulators of G1-phase progression[J].Genes Dev,1999,13(12):1501-1512.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
