光皮樹的組織培養研究初探
2011-06-11 02:12:12彭曉英劉永濤周雙德趙黎明張良波
湖南林業科技 2011年6期
關鍵詞:生長
劉 清,彭曉英*,劉永濤,周雙德,趙黎明,張良波
(1.湖南農業大學生物科學技術學院,湖南長沙 410128; 2.湖南省林業科學院,湖南長沙 410004)
光皮樹 (Swida wilsoniana Wanaer)為山茱萸科(Cornaceae)梾木屬 (Swide opiz)落葉灌木或喬木,其干果含油率達26%~36%,且果實油中含油酸和亞油酸高達77.68%,可通過酯化工藝制取生物柴油[1-2]。其燃料特性和動力性與0#柴油相似,是一種優良的代用燃料[3-5]。對石油能源枯竭的擔憂,以及利用光皮樹油制取生物柴油研究的快速開展,使得光皮樹作為重要的生物柴油原料得到廣泛關注。
目前,光皮樹的繁殖主要是通過播種育苗、扦插育苗和嫁接繁殖[6-8],播種育苗子代個體分化明顯,難以保持母本的優良性狀;而扦插和嫁接育苗周期較長,易受病蟲害影響。如能運用組織培養技術建立光皮樹無性系,則可以不受氣候影響,在較短時間內繁育大量光皮樹良種苗木[7]。因此,本研究擬采用組織培養手段,通過愈傷組織的誘導和分化及腋芽增殖等途徑,旨在建立光皮樹離體繁殖體系。
1 材料與方法
1.1 材料
采自湖南省林業科學院經濟林苗圃的光皮樹成熟種子 (2007年11月成熟)和嫩枝條 (2008年3月采集)。
1.2 方法
1.2.1 光皮樹種子和嫩枝條處理
(1)光皮樹種子處理:將光皮樹種子摩擦破皮,消毒后接種于基本培養基中。15 d左右種子開始萌動,長出子葉和胚軸。分別以子葉和胚軸為外植體進行培養。
(2)光皮樹嫩枝條處理:將采集的嫩枝條放在裝有少許自來水的燒杯中,培育一周左右。待光皮樹腋芽伸長后,分別以莖段、頂芽、葉片和帶腋芽的莖段為外植體進行培養。
1.2.2 培養條件 培養室溫度25±2℃,日光燈光照14 h/d,光照強度為2 000 lx。
1.2.3 取材與接種 將光皮樹種子接種到基本培養基上,獲得無菌種子苗。①切割其胚軸和子葉分別以平放和豎插兩種方式轉入愈傷組織誘導培養基中培養。②切割莖段、頂芽、葉片和帶腋芽莖段為外植體,莖段和頂芽切段、葉片切塊,用70%酒精快速浸泡,無菌水沖洗后轉入0.1%的升汞中消毒,無菌水沖洗3次。70%酒精的浸泡時間取0、30、60 s,0.1%升汞溶液的消毒時間取4、6、9 min。然后接種于基本培養基上進行培養。10 d后統計污染率,20 d后統計褐化率。
1.2.4 愈傷組織誘導 將切割的莖段、頂芽、葉片、胚軸、子葉和帶腋芽的莖段分別接種于附加不同濃度配比的2,4—D和KT的MS培養基中,在愈傷組織誘導試驗過程中,2,4—D設3個濃度梯度,KT設3個濃度梯度。出愈率為培養30 d后誘導出愈傷的外植體數與接種外植體數之比。
1.2.5 不定芽的誘導 將誘導出的愈傷組織切塊,接種于附加不同濃度配比的6—BA和NAA的MS培養基上進行培養,觀察愈傷組織的生長情況。愈傷組織的誘導分化試驗設計中6—BA設5個濃度梯度,NAA設2個濃度梯度。分化率為培養30 d后分化的愈傷組織數與接種愈傷組織數之比。
1.2.6 腋芽增殖 將經過處理的腋芽接種于添加不同濃度配比的6—BA和KT的MS培養基上進行培養,觀察腋芽增殖過程中所發生的變化。其中帶腋芽的莖段分為平放和豎插兩種方式。在試驗過程中,6—BA設5個濃度梯度,KT設2個濃度梯度。增殖率為培養30d后總的腋芽數目與最初接種時的腋芽數目的比值。
1.2.7 根的誘導 待叢芽長至2~3 cm時,切割分離,轉入不同濃度配比的IAA和NAA的MS培養基上,誘導生根。NAA和IAA均設3個濃度梯度。
2 結果與分析
2.1 不同消毒時間對不同外植體的影響
用70%酒精和0.1%升汞溶液對光皮樹的莖段、頂芽、葉片分別進行消毒處理,不同處理時間的效果見表1。
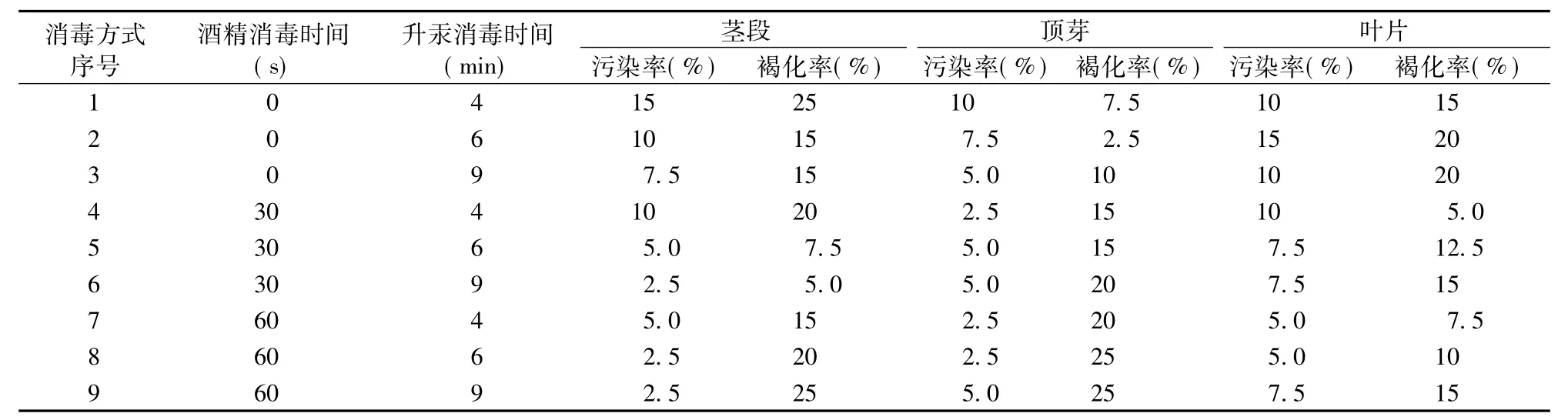
表1 不同消毒試劑和消毒時間對外植體的影響Tab.1 Effects of different sterilization reagent and sterilization time on explants
表1結果顯示:單獨使用升汞消毒時,各外植體的污染率最高,如莖段污染率達到15%,而與酒精配合使用后,污染率有明顯下降,當酒精中浸泡時間為30 s時,莖段污染率下降至10%。這是因為光皮樹莖段、頂芽和葉片表面都有細柔毛,而70%的酒精穿透力較強,能短時間滲透消毒,故與升汞搭配使用較為適宜。隨酒精和升汞溶液中消毒時間的延長,外植體的污染率呈下降趨勢,而褐化漸趨明顯,褐化率上升。從表1中結果看出,莖段作為外植體時,在酒精中浸泡30 s后清洗,再于升汞溶液中消毒4~6 min時,材料的污染率較小,且褐化率相對較低,最佳的消毒時間為70%酒精30 s,0.1%升汞9 min;頂芽最佳的消毒時間為70%酒精30 s,0.1%升汞4 min;而葉片表面柔毛較多,故70%酒精60 s后0.1%升汞中處理4 min,能達到較理想的效果。
2.2 不同生長調節劑配比對不同外植體愈傷組織誘導的影響
光皮樹愈傷組織誘導頻率與外植體來源有直接關系。六種外植體在接種到不同生長調節劑配比的培養基中,均能誘導出愈傷組織,但誘導條件和愈傷組織的生長情況不盡相同。(見表2)葉片在第8天開始卷曲,切口邊緣開始長出亮白色的愈傷組織 (圖1a);莖段在第4天開始啟動 (圖1b),10 d后長出淡黃色致密愈傷組織,生長迅速 (圖1c);頂芽啟動稍晚些,但生長較快,后期長勢良好 (圖1g)。
結果表明:2,4—D濃度在1~4 mg/L范圍內隨著濃度的增加,愈傷組織的誘導率呈上升趨勢,但2,4—D濃度過高 (4 mg/L)抑制愈傷組織的產生,導致愈傷組織出現玻璃化現象。當2,4—D和KT濃度分別為2 mg/L和0.5 mg/L時,愈傷組織的誘導率較高。其中莖段在最適培養基上出愈率最高,達到90%,愈傷組織質地致密顏色淡黃綠色,生長速度較快 (圖1c);葉片的出愈率次之,后期達到89%,愈傷組織質地疏松顏色發白 (圖1a);子葉的出愈率達到88%,形成了質地疏松,淡黃色愈傷組織,后期呈現玫瑰紅色(圖1d);帶腋芽的莖段出愈率達到84%,形成了疏松,玻璃化的愈傷組織 (圖1e);胚軸的出愈率達到81%,形成了疏松玻璃化的愈傷組織 (圖1f);頂芽的出愈率也達到85%,形成了致密,淡黃色的愈傷組織(圖1g)。
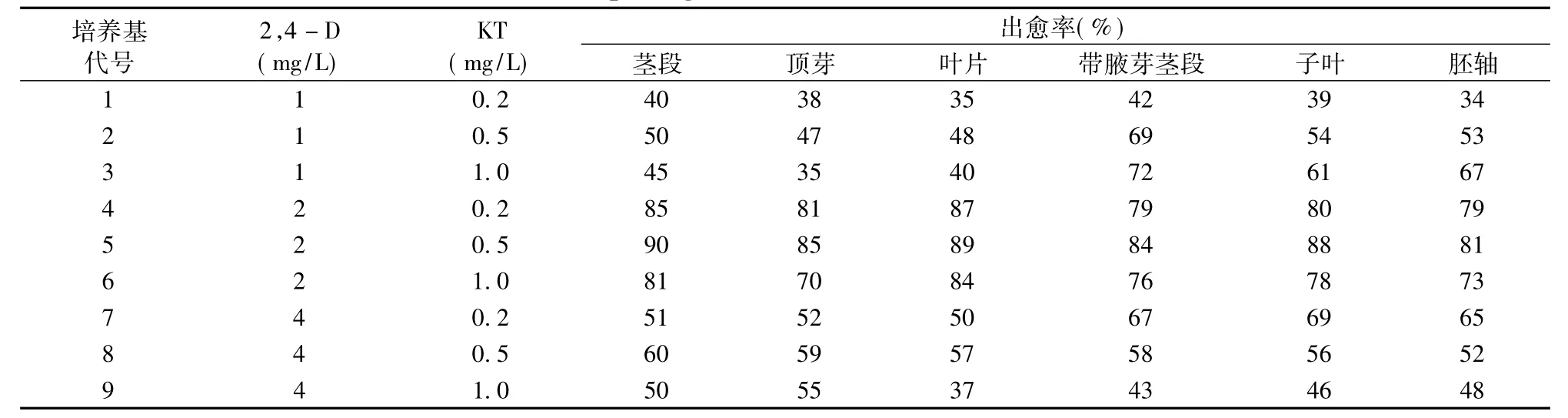
表2 不同濃度組合植物生長調節劑對愈傷組織誘導的影響Tab.2 Effects of different plant growth substances combination on callus induction

圖1 光皮樹不同外植體的愈傷組織 (一)Fig.1 The callus of different explant of S.wilsoniana(a)
2.3 不同生長調節劑濃度組合對愈傷組織分化的影響
將切塊的愈傷組織轉接到分化培養基 (MS+6BA 2.0 mg/L+NAA 0.2 mg/L和MS+6BA 4.0 mg/L+NAA 0.4 mg/L)中進行培養,20 d后發現由葉片、胚軸、子葉和帶腋芽的莖段長出的愈傷組織均呈現玻璃化和黏液化現象。而由莖段長出的愈傷組織質地依然致密,顏色淡黃綠色。因此,將由莖段誘導出的愈傷組織轉入不同生長調節劑濃度配比的培養基上,35 d后統計愈傷組織上的不定芽分化率并記錄愈傷組織的變化情況。結果見表3。
實驗結果表明:由莖段誘導出的愈傷組織接種于上述供試培養基中均未分化。隨著6—BA濃度的升高,愈傷組織的質地由疏松變化成稍致密,顏色由透明變為淺黃色再到淺黃綠色,為分化提供了可能性。6—BA(4.0 mg/L)和NAA(0.5 mg/L)稍有利于致密淺黃綠色愈傷組織的繼代培養。
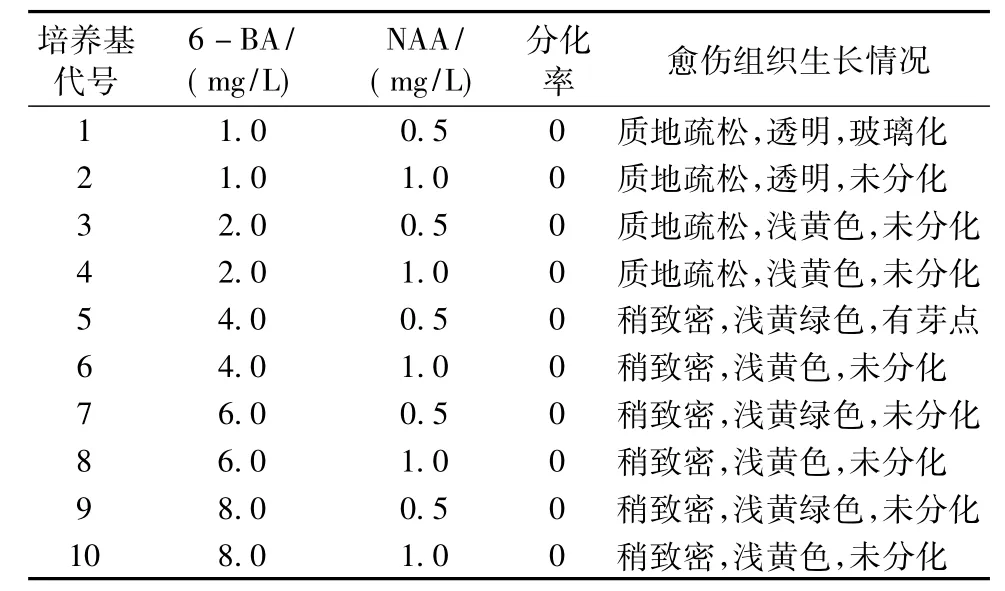
表3 不同生長調節劑濃度組合對不定芽分化的影響Tab.3 The concentration of different growth regulator combinations on the differentiation of adventitious buds
2.4 不同生長調節劑濃度組合對光皮樹腋芽增殖的影響
將光皮樹帶腋芽的莖段接種于不同生長調節劑濃度配比的培養基中,接種方式分為平放和豎插。觀察腋芽增殖的情況,并記錄。結果見表4。
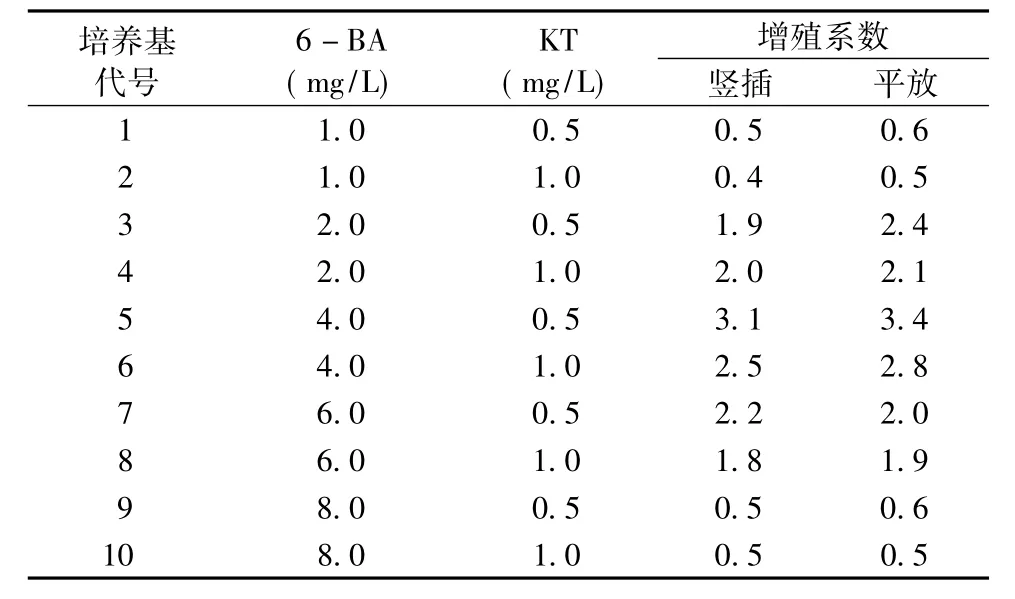
表4 不同生長調節劑濃度配比對腋芽增殖的影響Tab.4 Different growth regulators and their concentration ratio on the impact of axillary bud proliferation
表4結果表明:帶腋芽的莖段在進行平放和豎插時,二者的增殖系數相差無幾。6—BA的濃度處于1.0 mg/L和8.0 mg/L時,腋芽的增殖系數較低。從總體上來看,隨著6—BA濃度的增大,增殖系數先增后降。6—BA濃度為4.0 mg/L、KT為0.5 mg/L時,兩種接種方式的增殖系數均最高,平放時可達3.4。帶腋芽的莖段平放時,接種4 d后腋芽開始萌動,呈淺嫩綠色且莖段基部膨大,10 d后在腋芽基部周圍產生數量不等的叢芽數 (2~6個)(圖2a);豎插時,接種6 d后腋芽開始萌動,15 d后產生數量不等的叢芽數 (1~5個)。其中平放時莖段腋芽增殖效果較好,生長明顯,長勢旺盛,莖段健壯 (圖2c)。

圖2 光皮樹不同外植體的愈傷組織 (二)Fig.2 The callus of different explant of S.wilsoniana(b)
2.5 不同生長調節劑濃度組合對生根的影響
將叢芽轉接入供試生根培養基中,叢芽基部逐漸褐化或未見根的分化。其原因可能是因為光皮樹為多年生木本植物,而多年生木本植物普遍存在生根較困難或生根時間較長等現象[9]。另外供試生長調節劑的濃度組合可能不適宜光皮樹的生根培養。
3 討論
3.1 子葉和胚軸接種方式對愈傷組織誘導的影響
在本試驗中發現,子葉和胚軸誘導愈傷過程中,子葉和胚軸平放對誘導愈傷較好,這可能是因為子葉和胚軸平放,能充分接觸培養基,從中吸收養分,接受生長調節劑的誘導,因此兩端切口處的出愈率高,愈傷組織長勢較好;當子葉和胚軸插入培養基,僅切面細胞接觸,吸收及接受生長調節劑誘導的面積小,向上運輸養分和生長調節劑困難,導致子葉和胚軸出愈率下降。在子葉和胚軸誘導不定芽再生過程中,僅僅產生愈傷,均未誘導出芽點,可能是細胞分裂素和生長素的比值影響了芽的分化,需要進一步探索誘導分化的培養基。
3.2 外植體類型與愈傷組織誘導的關系
來源于光皮樹不同部位的愈傷組織在結構和顏色及生長特性等方面都有明顯的差異。本試驗以葉片、莖段、頂芽、子葉、胚軸和帶腋芽的莖段為外植體進行愈傷組織的誘導,該愈傷組織質地疏松,在添加較高濃度的6—BA與適量NAA的誘導培養基中,以莖段為外植體時,能在切口處膨大產生質地稍硬的致密愈傷組織,顏色變成淺黃綠色。在同樣的培養條件下,葉片、頂芽、子葉和胚軸等外植體卻只能誘導出愈傷組織,顏色一般為透明,質地疏松。分析其原因,可能是不同部位外植體的組織結構和細胞類型不同,它們產生愈傷組織所需要的生長調節物質種類和所需量也有很大的差別。故以莖段為外植體,愈傷組織的啟動能力明顯比其他外植體強,誘導時間短于其他外植體的誘導時間,且愈傷組織生長速率快,細胞分裂時間短,這與有關報道所述光皮樹植株幼嫩莖段細胞分裂旺盛,易于再分化[10]相符。
3.3 不同植物生長調節劑對外植體愈傷組織誘導和分化的影響
植物組織培養的愈傷組織分化過程是生長素含量逐步降低、細胞分裂素含量逐步升高的過程。本實驗中愈傷組織未能分化,一種可能是由于還不了解光皮樹自身激素的狀況,沒有找到適合誘導分化的細胞分裂素與生長素比值;另外一種可能是由于高質量濃度的2,4—D對外植體及愈傷組織的傷害。一些研究者認為,盡管較高質量濃度的2,4-D有利于胚性愈傷組織的誘導,但高質量濃度的2,4—D使整個培養系統的生長調節物質比例始終處于高比例的生長素類似物狀態,從而不利于隨后進行的愈傷組織分化再生芽的過程[11-12];第三種可能是由于所誘導的愈傷組織屬于非胚性愈傷組織,這與有關資料所述的“非胚性愈傷組織分化率低,甚至不分化”的觀點相符合[12-13]。
4 結語
本文分別從光皮樹愈傷組織的誘導、增殖、分化和腋芽增殖等途徑進行研究,各外植體均能誘導愈傷組織,但不定芽分化困難,在愈傷組織誘導不定芽的途徑中,愈傷組織在分化培養基上相當長一段時間內形態上幾乎沒有變化。有研究者認為初代培育時培養基配方中的酸性物質使外植體處于酸性環境中并導致植物正常細胞首先發生細胞壁酸性降解,隨后出現原生質體脫離細胞壁,進一步發生細胞器重組或細胞重建[14]。在愈傷組織形成后,如果轉移過早,還未形成成熟的脫分化組織;過晚,愈傷組織可能褐化或喪失進一步分化的能力。所以探索出合適愈傷組織的轉移及其分化時刻也是間接誘導不定芽途徑中非常重要的工作環節。今后還應在繼代培養、不同時期的愈傷組織細胞切片以及次生代謝物方面作深入的研究,從而在生理的基礎上,了解其生長發育的機制。特別是如能從分子水平上檢測光皮樹不同發育階段對各種理化因子的需求,以及通過探索光皮樹不同生長期對各種生長調節劑的應答機制,了解不同生長調節劑之間和光皮樹的相互作用,從而建立一套完整的離體培養體系。這將對實現光皮樹工廠化育苗及進一步利用基因工程技術實現對高油含量光皮樹遺傳轉化奠定技術基礎,具有重要實踐意義。
[1]萬志洲,黃利斌,等.明孝陵光皮樹的生物學特性及繁育技術研究[J].林業實用技術,2007(2):3-5.
[2]梁仰貞.值得發展的油料植物——光皮樹 [J].植物雜志,1996(2):12.
[3]梁仰貞.光皮樹栽培技術 [J].特種經濟植物,2007,10(3):38-39.
[4]李昌珠,蔣麗娟,程樹棋.生物柴油研究現狀與商業化應用前景[C] //中國生物質能技術研討會論文.南京,2002.
[5]李正茂,鄧新華,李黨訓.光皮樹經濟性狀及生物質液體燃料開發研究構想[J].湖南林業科技,1996,23(2):11-13.
[6]成訓妍.光皮樹是珍貴的木本食用油料資源 [J].生物與特產,1990(6):28.
[7]本瑪麗,王曉明,李永欣,等.光皮樹優良無性系組織培養的無菌體系建立[J].湖南林業科技,2010,37(2):5-8.
[8]曾紅燕,李昌珠,蔣麗娟,等.不同方法提取光皮樹籽油的GC-MS分析 [J].中國生物工程,2004,11(24):83-86.
[9]朱玉球,童再康,黃華宏,等.紅葉石楠硬枝水培生根試驗[J].浙江林學院學報,2004,21(1):28-32.
[10]許智宏,劉春明.植物發育的分子機理 [M].北京:北京科技出版社,1999.
[11]蔣澤平,梁珍海,吳綱,等.秤錘樹的組織培養和快速繁殖[J].植物生理學通訊,2005,41(2):191.
[12]潘國才,婁漢平,吳麗敏,等.遼東楤木組織培養快繁技術[J].現代農業,2009(4):70-71.
[13]張小紅,陳彥生,康冰,等.激素對香椿腋芽增殖生長的效應 [J].西北植物學報,2001,21(4):756-760.
[14]祖元剛,于景華,唐中華,等.植物葉片愈傷組織形成的可能機制 [J].植物研究,2005,25(1):1-2.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
