埃及吹綿蚧在園林植物上的空間分布型研究
2011-06-09 05:57:26柴芳華安新城
環境昆蟲學報 2011年4期
關鍵詞:植物
柴芳華,郭 強,蔣 露,安新城
(1.中國科學院華南植物園,廣州 510520;2.深圳野生動植物保護管理處,深圳 518001;3.廣東省昆蟲研究所,廣州 510260)
埃及吹綿蚧Icerya aegyptiaca(Douglas),隸屬于同翅目 Homoptera,珠蚧科 Margarodidae,吹綿蚧屬Icerya,是一種重要的的園林害蟲,在我國大部分地區都有分布 (王子清,1982)。該蟲食性雜,寄主植物多達數百種,特別喜食木蘭科植物(劉東明等,2003)。埃及吹綿蚧主要以雌成蟲和若蟲危害植物的葉芽、嫩枝及枝條,成群聚集在葉背面或嫩枝上吸食植物汁液,受害樹木葉色發黃,枝梢枯萎,引起落葉落果,樹勢衰弱,甚至枝條或全株枯死,同時,排泄蜜露,誘致煤污病,影響植物光合作用 (徐公天,2003)。近年來在華南地區,埃及吹綿蚧在城市綠化植物上的為害呈上升趨勢 (岑炳沾和蘇星,2003),為了有效、安全、持續地控制埃及吹綿蚧的危害,深入研究埃及吹綿蚧在園林植物上的生物學和生態學特性是非常必要的。
本文對埃及吹綿蚧的空間分布型進行了較為細致的研究,并探討了埃及吹綿蚧的種群擴散和分布規律,以期為埃及吹綿蚧的防治提供有益的基礎信息。
1 材料與方法
調查地點:深圳筆架山公園,植被以草本和灌木為主,調查時間為2010年8月。
調查方法:在埃及吹綿蚧發生區隨機選取79株被害植物,每株植物分東西南北4個方向,每個方向隨機選取10個葉片,記錄每片葉上的埃及吹綿蚧數量,共計3160個樣本。
數據處理:運用統計軟件SPSS 13.0和EXCEL 2002版本,對樣本進行頻度分析,并分別擬合泊松分布、奈曼分布和負二項分布,采用卡方檢驗其擬合度。
泊松分布的概率模型:

其中,Px是一個樣本中出現x頭昆蟲的概率;N是樣本數;m是每個樣本中個體數的平均數;e是自然對數的底。
奈曼分布的概率模型:
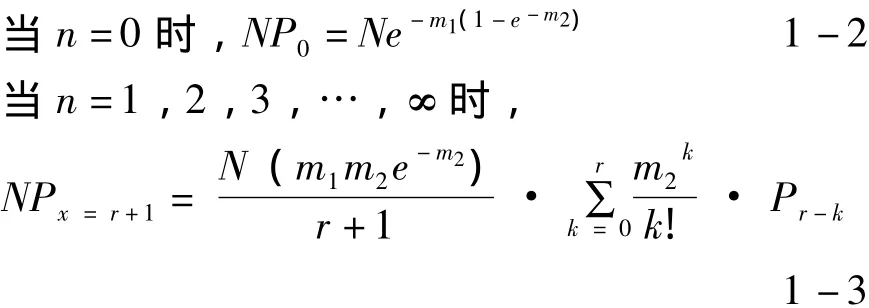
其中,P為樣本中出現n頭昆蟲的概率;N是樣本數;m1、m2為估計參數,可通過方差 (V)和平均數求得,如下式:
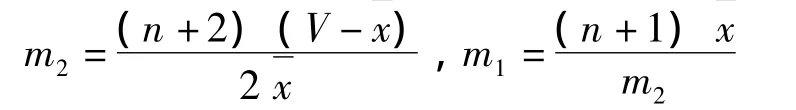
負二項分布的概率模型:
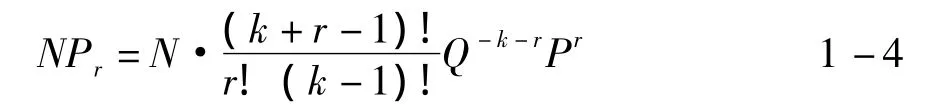
其中,Pr為樣本中出現r頭昆蟲的概率;N是樣本數;k、Q、P為估計參數,可通過方差 (V)和平均數求得,如下式:

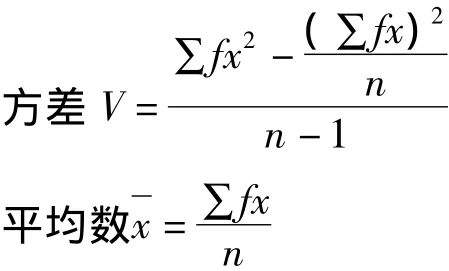
其中:x為蟲數,f為頻數,n為樣本總數。卡方檢驗公式:

其中:x為實際值,c為理論值。
2 結果與分析
調查了79株植物,每株植物4個方向,每個方向10個樣本,對原始數據進行頻度分析,結果如表1。

表1 埃及吹綿蚧空間分布的頻度分析Table 1 Frequency analysis of spatial distribution for Icerya aegyptiaca
從表1可以看出,樣本中大于10頭蟲數的頻率非常小,其累積百分率小于1%,為了簡化計算,可予以省略。
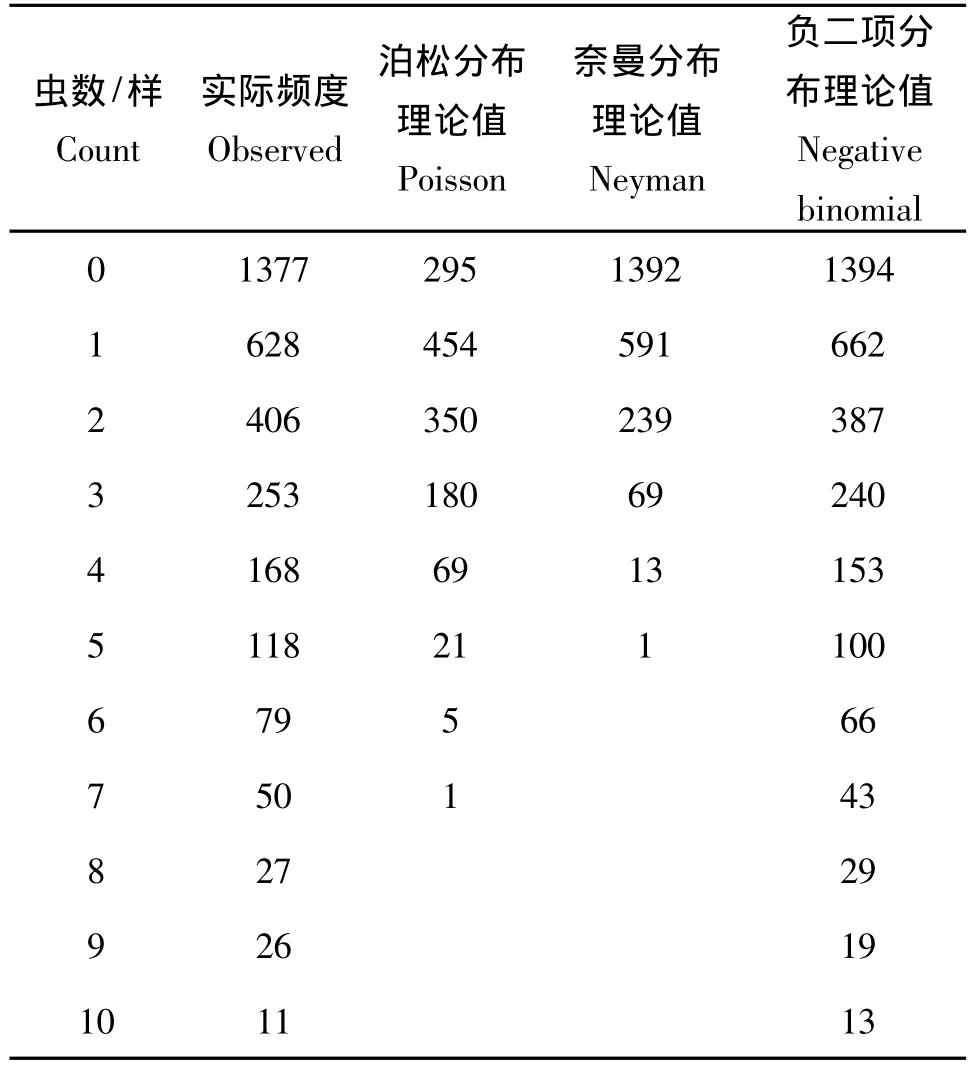
表2 埃及吹綿蚧的實際頻度與三種理論頻度的比較Table 2 Compared with observed and predicted data on 3 spatial patterns for Icerya aegyptiaca
泊松分布為隨機分布,通過平均值計算各理論值,從表中可以看出理論值與實際頻度相差較遠。
奈曼分布為核心分布,其分布的理論值根據n的變化而有很大差異,表2中的理論值是n→∞時的奈曼分布理論值,因n=0和n=2時的理論值與實際頻度相差甚遠,故未在表中列出。
負二項分布為聚集分布,其k、Q、P參數可由樣本信息來估算,其中k值的估算最為關鍵,由于k值的估算方法不同,可導致理論值產生很大的變化。本文采用了較為精確的零頻率法,其一是因為k值小于1,其二是樣本數據完全符合零頻率法要求的兩個條件:

從表2中可以看出,用較為精確的k值計算出的負二項分布理論值與實際頻度有很好的吻合,泊松分布、奈曼分布和負二項分布的擬合度檢驗見表3。
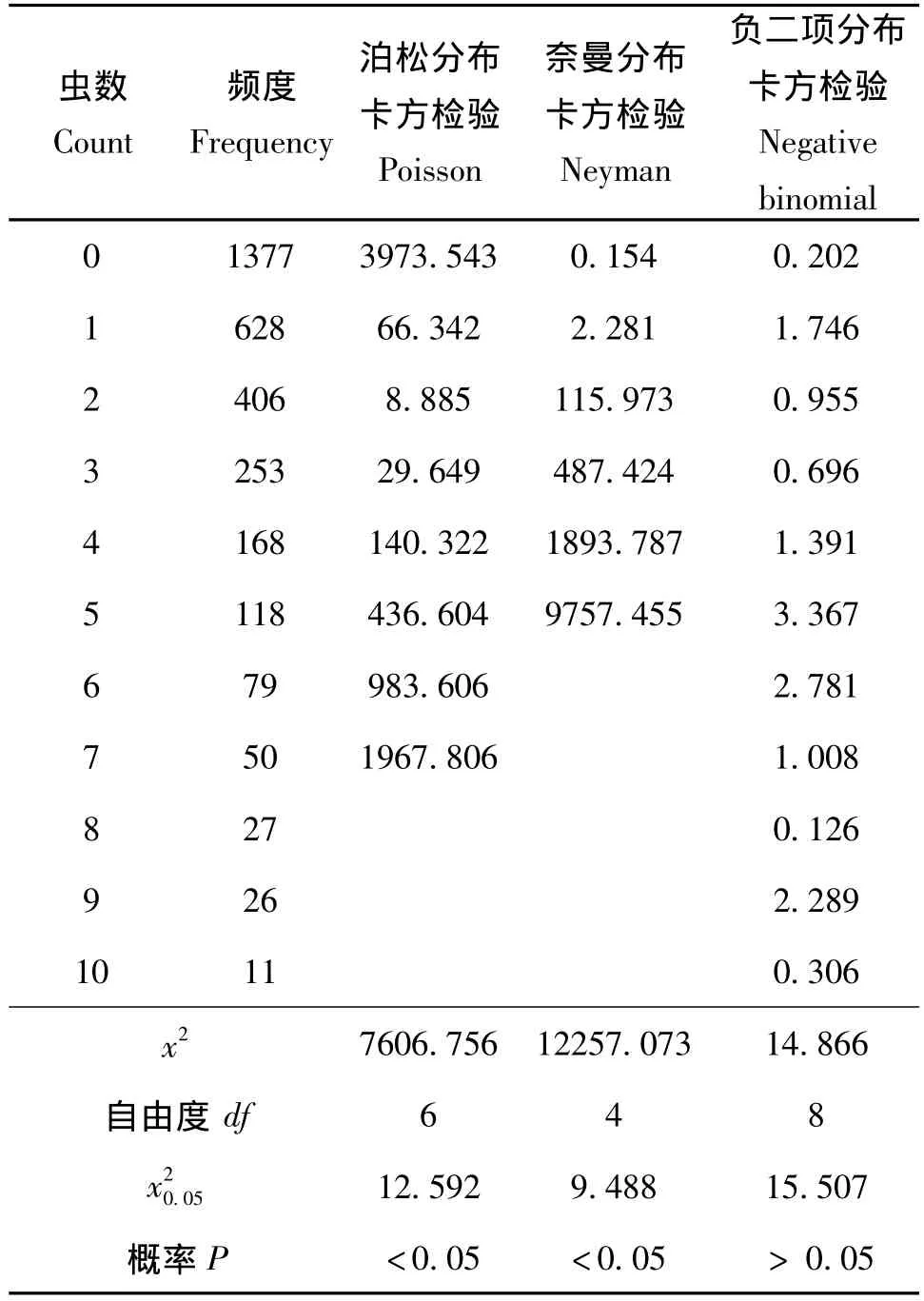
表3 埃及吹綿蚧三種分布型的卡方檢驗Table 3 Chi square test of 3 spatial patterns for Icerya aegyptiaca
從表3的卡方檢驗表中可以看出,埃及吹綿蚧的頻度數據極不適合泊松分布和奈曼分布 (P<0.05),而與負二項分布擬合得較好 (P>0.05)。
3 結論與討論
自然狀態下的昆蟲種群在田間的空間分布受多種因素疊加的影響,寄主植物的分布、天敵、溫濕度、光照和固體障礙物等都不同程度地導致昆蟲種群在空間上呈多種變化的分布 (Andrew et al.,2003;Sasha et al.,2009),在深圳筆架山調查區域內,植物種類以灌木和小喬木為主,由于人類活動的頻繁干擾,林下植被較少,昆蟲多樣性不高,因此預期埃及吹綿蚧的擴散、遷移和分布,主要受氣候、寄主植物和自身生物學特性的影響。
從本文的結果來看,埃及吹綿蚧在野外極少出現隨機分布的狀況,這與大多數固定取食的昆蟲相類似,隨著食物資源的斑塊狀分布,不擅活動的昆蟲也相應地表現為斑塊狀分布 (Jeger,1999)。
奈曼分布是不均衡分布,即昆蟲在棲息地呈多數小集團,形成核心,核心大小約相等,并自核心作放射狀蔓延。從表2可以看出,除蟲數為0和1的情況外,埃及吹綿蚧也極不適合奈曼分布,但是,奈曼分布與負二項分布之間是一個連續變化的過程,因此不排除埃及吹綿蚧在發生初期,會趨向于符合奈曼分布,隨著種群數量的增加、分布范圍的擴大,再由奈曼分布轉化為核心大小不等的分布 (如波利亞分布),以及多個核心融合而成的疏密相間的分布 (負二項分布)。
本文中埃及吹綿蚧的種群分布體現為典型的聚集分布,并且每個樣本的平均蟲數較少(=1.54頭),這說明埃及吹綿蚧單位面積內種群密度較低,處于盛發期而又密度較低,這種現象暗示埃及吹綿蚧可能具有較快的擴散速度,結合埃及吹綿蚧繁殖力強、寄主范圍廣的特性,這無疑給防治帶來了一定的難度。
References)
Andrew WF,Zdis IK,Barbara WS,Suzanne JC,Moira AM,Joe NP,Ingrid HW,2003.Spatial distribution of pest insects in oilseed rape:implication for integrated pest management.Agriculture,Ecosystems and Environment,95(2):509 -521.
Chen BZ,Su X,2003.Control of Pest and Disease on Ornamental Plants.Guangzhou:Science and Technology Publishing of Guangdong.68-69.[岑炳沾,蘇星,2003.景觀植物病蟲害防治.廣州:廣東科技出版社.68-69]
Jeger MJ,1999.Improved understanding of dispersal in crop pest and disease management:current status and future direction.Agricultural and Forest Meteorology,97(4):331-349.
Liu DM,Chen HF,Yi QF,Zen QW,Xing FW,2003.Damage and control of Icerya aegyptiaca on Magnoliaceae plants.Plant Protection,29(6):36-38.[劉東明,陳紅鋒,易綺斐,曾慶文,邢福武,2003.埃及吹綿蚧在木蘭科植物上的發生危害及防治.植物保護,29(6):36-38]
Sasha CV,Helen S,Ian RD,2009.Annual and seasonal patterns of insect succession on decomposing remains at two locations in Western Australia.Forensic Science International,193(1):26 -36.
Wang ZQ,1982.Scale Insect of Agriculture Area in China.Beijing:Agriculture Press,31-32.[王子清,1982.中國農區的介殼蟲.北京:農業出版社,31-32]
Xu GT,2002.Current situation and countermeasure for pest and disease on urban ornamental plant in China.Forest Pest and Disease,12(1):48-52.[徐公天,2002.我國城市園林植物病蟲害的現狀及對策.中國森林病蟲,12(1):48-52]
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
