茄鏈格孢菌侵染馬鈴薯葉片過程的細胞學觀察
2011-05-22 03:55:12左豫虎韓青梅康振生臺蓮梅
中國蔬菜 2011年14期
左豫虎 史 潔 韓青梅 康振生 臺蓮梅 劉 銅
(1黑龍江八一農墾大學,黑龍江 大慶 163319;2西北農林科技大學植物保護學院,陜西省農業分子生物學重點實驗室,陜西 楊凌 712100)
馬鈴薯早疫病(Potato early blight)是一種由茄鏈格孢菌〔Alternaria solani(Ell.et Mart.)Jones et Grout〕引起的對馬鈴薯危害比較嚴重的病害,可以導致馬鈴薯產區大面積減產,帶來巨大的經濟損失(Pelletier & Fry,1990)。該病在我國各大馬鈴薯產區均有不同程度發生,局部地區其危害程度不亞于晚疫病(楊殿賢和苑風瑞,2007)。利用組織病理學和電鏡技術研究茄鏈格孢菌與馬鈴薯互作的組織病理學和超微結構特征,可為在細胞水平上闡明茄鏈格孢菌的致病機制及馬鈴薯的抗病機制提供理論依據。Dita等(2007)對馬鈴薯早疫病病菌侵染過程進行了研究,發現茄鏈格孢菌可從表皮細胞的相鄰部位直接侵入,沒有觀察到從氣孔入侵,并且發現侵染與品種抗性無關,但侵染后抗病品種較感病品種表現出明顯的過敏性壞死反應。而在我國鮮見茄鏈格孢菌與馬鈴薯互作的組織病理學和細胞學研究的相關報道。本試驗采用掃描和透射電鏡技術系統研究了茄鏈格孢菌在抗、感早疫病的馬鈴薯離體葉片上的侵染過程及超微結構特征,旨在為從細胞水平上闡明茄鏈格孢菌的致病機制及馬鈴薯的抗病機制提供理論依據。
1 材料與方法
1.1 供試品種
供試馬鈴薯品種為東農303(感病)和克新1號(抗病),由黑龍江省克山農場馬鈴薯研發中心提供。品種抗性由黑龍江八一農墾大學植物病理研究室接種鑒定(臺蓮梅 等,2010)。
1.2 供試菌株
供試茄鏈格孢菌菌株SH0806為2008年分離自黑龍江省綏化市采集的馬鈴薯品種富津(遼寧省本溪市馬鈴薯研究所培育)的早疫病病葉,由本大學植物免疫研究室提供。。
1.3 分生孢子懸浮液的制備
試驗于2009年10月~2010年3月在西北農林科技大學陜西省農業分子生物學重點實驗室進行。用打孔器打取PDA平板上生長5~7 d、直徑為5 mm的茄鏈格孢菌菌碟,接入水瓊脂平板,25 ℃黑暗培養3 d后產生大量分生孢子,加少量無菌水輕輕震動,使分生孢子能夠充分的散落于水中。用血球計數板將分生孢子懸浮液的濃度調整到1.0×105個·mL-1備用。
1.4 離體葉片接種方法
選擇3~4葉期的馬鈴薯同一部位葉片,取1.3制備的孢子懸浮液10 μL,滴于葉片背面,用于透射電子顯微鏡取樣;取孢子懸浮液10 μL滴于葉片正面,用于掃描電子顯微鏡取樣,以蒸餾水接種為對照。置于放有1~2張濕潤濾紙的培養皿內,25 ℃恒溫培養箱中黑暗保濕培養。
1.5 掃描電鏡(SEM)樣品的制備與觀察
分別于接種后2、4、6、8、10 h取樣,將接種部位的葉片切成1 cm2的小塊,按康振生(1995)的方法制備樣品。葉樣經抽真空后置于4 %的戊二醛磷酸緩沖液(0.1 mol·L-1、pH 6.8)中4 ℃固定過夜,然后用相同濃度磷酸緩沖液(PBS)沖洗5~6次,每次20 min,用30 %、50 %、70 %、80 %、90 %、100 %乙醇脫水,每次30 min,再用100 %丙酮脫水2次,每次30 min,最后用醋酸異戊脂置換2次,每次30 min,二氧化碳臨界點干燥、粘樣、噴金后在JEOL-JSM-6360LV型掃描電鏡下觀察、拍照。
1.6 透射電鏡(TEM)樣品的制備與觀察
分別于接種后12、24、48、72 h取樣,將接種部位的葉片切成0.3~0.5 cm2的小塊,按康振生(1995)的方法制備樣品。葉樣經抽真空后置于4 %戊二醛磷酸緩沖液(0.1 mol·L-1、pH 6.8)中4 ℃固定過夜,然后用相同濃度磷酸緩沖液(PBS)沖洗5~6次,每次20 min,于1 %鋨酸磷酸緩沖液(0.1 mol·L-1、pH 6.8)中固定2 h,用30 %、50 %、70 %、80 %、90 %、100 %乙醇脫水,每次30 min,再用100 %丙酮脫水2次,每次30 min,Epon812環氧樹脂包埋,在30 ℃和60 ℃下分別聚合24 h和48 h,最后用玻璃刀半薄切片定位,定位后用鉆石刀進行超薄切片。半薄切片于光學顯微鏡下觀察侵染點菌絲擴展情況并拍照,超薄切片經醋酸雙氧鈾—檸檬酸鉛雙重染色,于JEM-1230型透射電鏡下觀察、拍照。
2 結果與分析
2.1 分生孢子在葉片上的萌發
掃描電鏡觀察發現,感病品種東農 303葉片表面未萌發的茄鏈格孢菌分生孢子形態飽滿(圖版Ⅰ-1);在適宜培養條件下,接種2 h后,分生孢子開始萌發(圖版Ⅰ-2),分生孢子萌發時間在不同抗性品種葉片上無明顯差異(圖版Ⅰ-2、3、4);分生孢子可從孢子的多個位置(頭部、側面、尾部)萌發(圖版Ⅰ-2、3、4),萌發后孢子因營養物質的消耗而凹陷變癟(圖版Ⅰ-3、4);接種6 h后,抗病品種克新1號葉片上的菌絲沿葉片表面蔓延生長,有些菌絲產生了明顯的分泌物(圖版Ⅰ-5),而有些菌絲沒有產生分泌物(圖版Ⅰ-6)。
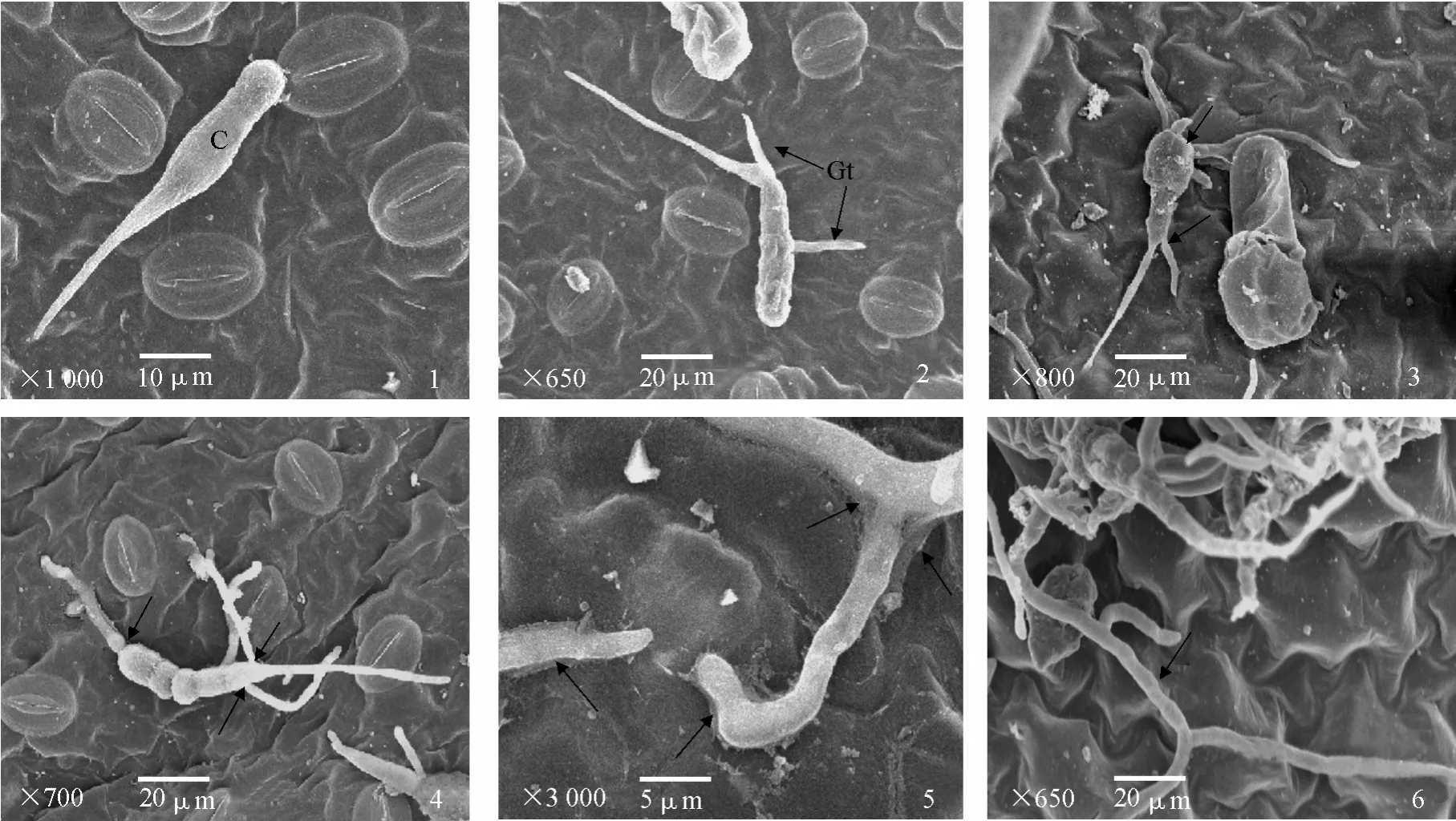
圖版Ⅰ 茄鏈格孢菌分生孢子在馬鈴薯葉片上萌發的掃描電鏡圖
2.2 分生孢子在葉片上的侵入
掃描電鏡觀察發現,接種 6 h后,馬鈴薯葉片表面茄鏈格孢菌菌絲大量蔓延,少數菌絲沿著寄主表皮細胞相鄰的凹陷處生長(圖版Ⅱ-1);隨著菌絲在寄主表皮細胞表面的擴展蔓延,菌絲頂端形成附著孢,部分菌絲越過氣孔蔓延生長(圖版Ⅱ-2),沒有觀察到通過氣孔侵入表皮細胞(圖版Ⅱ-2);接種8 h后,菌絲從表皮細胞相鄰的凹陷處直接侵入感病品種東農303葉片表皮細胞(圖版Ⅱ-3);接種10 h后,菌絲從表皮細胞相鄰的凹陷處直接侵入抗病品種克新1號的葉片表皮細胞(圖版Ⅱ-4)。說明茄鏈格孢菌侵入感病品種比侵入抗病品種早2 h。
2.3 菌絲在寄主體內的擴展
透射電鏡觀察發現,接種12 h后,茄鏈格孢菌菌絲在感病寄主細胞間隙生長(圖版Ⅲ-1);接種24 h后,菌絲穿過感病寄主細胞壁,存在于細胞壁和細胞膜之間,細胞膜變得不連續,可能是病原菌釋放出某些物質降解了寄主細胞膜(圖版Ⅲ-2),菌絲從一個細胞向另一個細胞生長(圖版Ⅲ-3);接種48 h后,大量菌絲存在于感病寄主細胞內和胞間(圖版Ⅲ-4)。
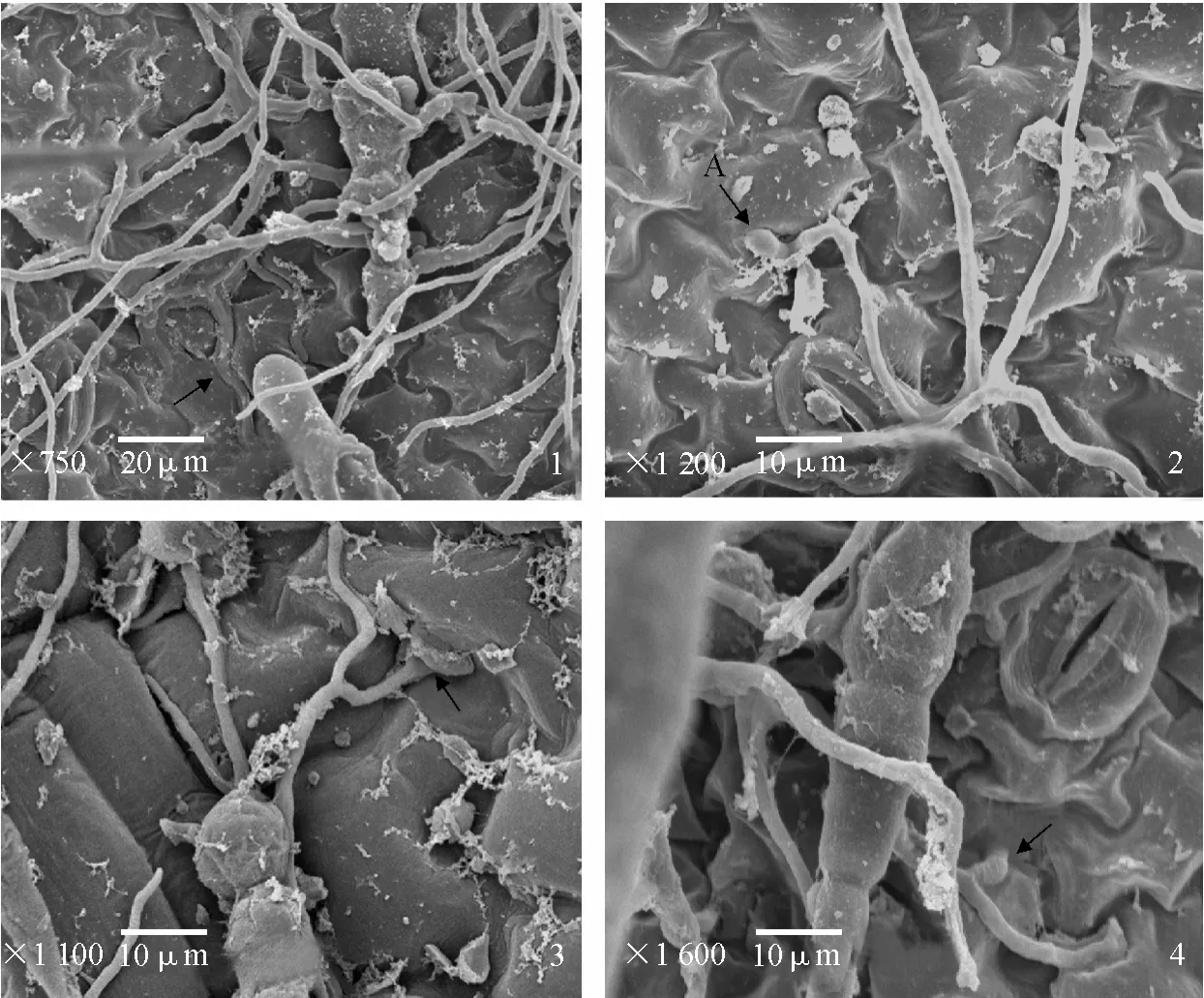
圖版Ⅱ 茄鏈格孢菌分生孢子在馬鈴薯葉片上萌發與侵入的掃描電鏡圖
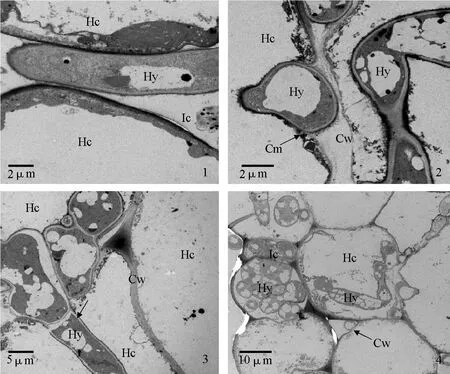
圖版Ⅲ 茄鏈格孢菌菌絲在寄主體內擴展的透射電鏡圖
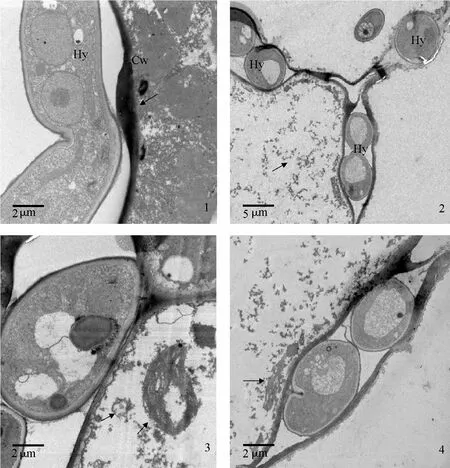
圖版Ⅳ 寄主細胞病理變化的透射電鏡圖

圖版Ⅴ 寄主細胞病理變化和茄鏈格孢菌菌絲在不同抗性品種葉片中的侵染情況
2.4 寄主組織的病理變化特征
接種24 h后,抗病品種克新1號寄主細胞壁明顯加厚(圖版Ⅳ-1),感病品種東農303未觀察到細胞壁加厚的現象,細胞內含物已基本全部消解(圖版Ⅳ-2);接種48 h后,抗病品種寄主細胞內含物出現不同程度的消解,還可見部分細胞器(圖版Ⅳ-3、4)。與感病品種接種24 h后寄主細胞內含物基本全部消解相比,抗病品種細胞消亡速度明顯慢于感病品種。
接種 72 h后,抗病品種細胞內含物基本全部消解(圖版Ⅴ-1),蒸餾水接種的對照寄主細胞內線粒體、葉綠體等各類細胞器均勻排列,清晰可見(圖版Ⅴ-2)。接種 48 h后,光學顯微鏡觀察半薄切片可見感病品種表皮細胞下侵染點存在大量菌絲(圖版Ⅴ-3),抗病品種表皮細胞下侵染點的菌絲(圖版Ⅴ-4)明顯少于感病品種。
3 結論與討論
利用光學顯微鏡、掃描和透射電子顯微鏡觀察茄鏈格孢菌侵染不同抗性馬鈴薯品種葉片的超微結構特征,發現接種2 h后,分生孢子開始萌發,分生孢子各個部位(頭部、側面、尾部)都能萌發;接種6 h后,菌絲頂端出現附著孢;接種8 h后,菌絲從表皮細胞間隙直接侵入感病品種表皮細胞內,接種10 h后才進入抗病品種表皮細胞內;接種24 h后,感病品種表皮細胞中侵染菌絲向相鄰細胞擴展,而此時抗病品種細胞壁出現明顯加厚的防衛反應,說明抗病品種對茄鏈格孢菌的侵染有一定的抗侵入和抗擴展能力。隨著茄鏈格孢菌在寄主細胞內的擴展,葉片發生了一系列的病理變化。接種24 h后,感病品種寄主細胞膜不連續,細胞內含物及各類細胞器已基本全部消解,抗病品種部分消解;接種48 h后,侵入抗、感病品種寄主表皮細胞內的菌絲數量差異明顯,感病品種菌絲量明顯多于抗病品種,抗病品種細胞內含物此時才基本全部消解。這表明抗病品種對茄鏈格孢菌的抗性機理存在抗菌絲擴展能力。
本試驗中分生孢子萌發、侵入時間明顯早于Dita等(2007)報道的接種6 h后分生孢子萌發,接種24 h后菌絲直接侵入感病品種,接種36 h后菌絲直接侵入抗病品種的試驗結果,這可能與供試菌株和寄主品種差異或與試驗條件有關,其原因有待于進一步研究。本試驗中發現抗病品種在接種24 h后出現細胞壁加厚的防衛反應,而感病品種未觀察到防衛反應現象。Dita等(2007)報道抗病品種的抗性表現是過敏性壞死反應。這是分別在細胞水平和宏觀水平對抗性表現的分析描述,并不矛盾。
病原菌的侵入方式一般分為 3種:傷口、氣孔、直接侵入。本試驗發現分生孢子萌發與品種抗性無關,部分菌絲在寄主表皮蔓延越過氣孔生長,沒有觀察到菌絲通過氣孔侵入寄主的現象,菌絲從表皮細胞間隙直接侵入,這與Dita等(2007)的研究結果一致。本試驗方法中,掃描電鏡樣品在葉片正面接種取樣,透射電鏡在葉片背面接種取樣。原因是:馬鈴薯屬雙子葉植物,葉片背面氣孔數和絨毛數明顯多于正面。正面接種掃描電鏡觀察,有利于避免絨毛茂密對觀察效果的影響;背面接種制備透射樣品,若病原菌能夠通過氣孔侵入,葉片背面氣孔數目多則利于茄鏈格孢菌的侵入。本試驗中還發現有些菌絲四周伴隨分泌物存在,該類分泌物的存在應該有利于分生孢子在寄主表面的附著與萌發,對其成分和作用有待于進一步研究證實。
康振生.1995.植物病原真菌的超微結構.北京:中國科學技術出版社:5-74.
臺蓮梅,梁偉伶,左豫虎,金光輝,靳學慧.2010.馬鈴薯不同品種感染早疫病菌后防御酶活性變化.植物生理學通訊,46(11):1147-1150.
楊殿賢,苑風瑞.2007.25 %嘧菌酯懸浮劑防治馬鈴薯早疫病田間藥效試驗.農藥科學與管理,28(8):28-29.
Dita M A,Brommonschenkel S H,Matsuoka K,Mizubuti E S G.2007.Histopathological study of the Alternaria solani infection process in potato cultivars with different levels of early blight resistance.Journal of Phytopathology,155(7-8):462-469.
Pelletier J R,Fry W E.1990.Characterization of resistance to early blight in three potato cultivars:receptivity.Phytopathology,80(4):361-366.
