斑茅種質資源形態(tài)性狀的變異研究
2011-04-25 10:01:18梁緒振鄢家俊白史且干友民
草業(yè)科學 2011年7期
關鍵詞:植物
梁緒振,鄢家俊,白史且,常 丹,干友民
(1.四川省草原科學研究院,四川 成都 611731; 2.四川農業(yè)大學草業(yè)科學系,四川 雅安 625014)
斑茅(Erianthusarundinaceum)又名大密、笆茅、大巴茅,系甘蔗(Saccharumofficinarum)的近緣植物,是蔗茅屬多年生、密叢高大草本。莖稈直立,高可達4 m,莖粗可達2 cm,具有分蘗力強、根系發(fā)達、抗旱性強、適應性廣泛、宿根性好等特性。其在丘陵坡地、河岸、路旁和礫石沙灘、荒地上生長良好,是優(yōu)良的固土護坡植物。在我國熱帶、亞熱帶至暖溫帶的廣大區(qū)域斑茅分布非常廣泛,因其莖稈和葉片纖維含量高,在民間作為編席、造紙的原材料及農村燃料的歷史由來已久[1-3]。斑茅在甘蔗育種中有較為特殊的育種價值和應用潛力,近年來國內外陸續(xù)開展了相關研究[4-6]。鑒于斑茅的生物學和生長特性,在我國南方大面積的荒山荒坡、鹽堿地、沿海灘涂地上種植這種禾草,既可作為能源草開發(fā),緩解能源供給緊張,又對改善我國生態(tài)環(huán)境具有重要意義,而且不會出現與傳統(tǒng)農業(yè)和畜牧業(yè)爭地的情況。
形態(tài)標記具有直觀有效、測量簡單等特點,是長期以來作物種植資源評價、育種后代選擇和遺傳多樣性研究最基本的標記。通過觀察形狀并結合一定的分析方法,就可以在短期內對所研究物種的遺傳變異水平有一個基本的認識[7-12]。本研究通過形態(tài)學方法對四川盆地及盆周山地60份野生斑茅材料進行分析,旨在檢測該地區(qū)斑茅遺傳多樣性狀況,了解其遺傳多樣性的分布格局,并為其開發(fā)利用與保護提供一定依據。
1 材料與方法
1.1試驗地概況 試驗地設在四川省草原科學研究院大邑縣韓場實驗基地,地處成都市大邑縣境內,地理位置30°25′ N,103°45′ E,海拔475 m,屬大陸性熱帶濕潤季風氣候。年平均氣溫15 ℃,最熱月(7月)平均氣溫26.1 ℃,最冷月(1月)平均氣溫5.5 ℃,極端最低氣溫-4.8 ℃,極端最高氣溫35.1 ℃,年降水量1 300 mm,全年日照時數1 033.8 h,年平均無霜期284 d。
1.2供試材料 供試材料為2008年8月采自四川盆地和盆周山地的野生斑茅。為了便于快速繁殖,以采集單株營養(yǎng)體為主。將采集的單株營養(yǎng)體移栽于塑料盆中,存活的材料為60株,以每株斑茅為1份材料,進行試驗田單株培養(yǎng),每份材料采樣地分布見表1。
1.3試驗設計 2010年3月將每份材料的所有分蘗分成單株,從中選出15個單株,移栽于小區(qū)內。小區(qū)面積為5 m×4 m,小區(qū)間隔1 m,將同份材料中多余的單株移栽到資源圃中,小區(qū)內單株的行距為1.5 m,株距為1.0 m,栽好后對其進行灌溉,以保證其成活率。成活的野生斑茅將作為室外指標的材料。
1.4觀測項目及測定方法
1.4.1營養(yǎng)器官性狀 葉片指標測定在初花期進行,每份種質選擇5~10株,10次重復。其中倒二葉片指從花序往下的第2片葉旗葉長、倒二葉長用卷尺測量;葉寬的測定部位為葉片的最寬處,用游標卡尺測量。莖粗測量部位為莖稈最粗處,用游標卡尺測量。株高和莖節(jié)數的觀測在乳熟期進行,用卷尺齊地測量植株垂直高度為株高,自生長點以上數其莖節(jié)數。
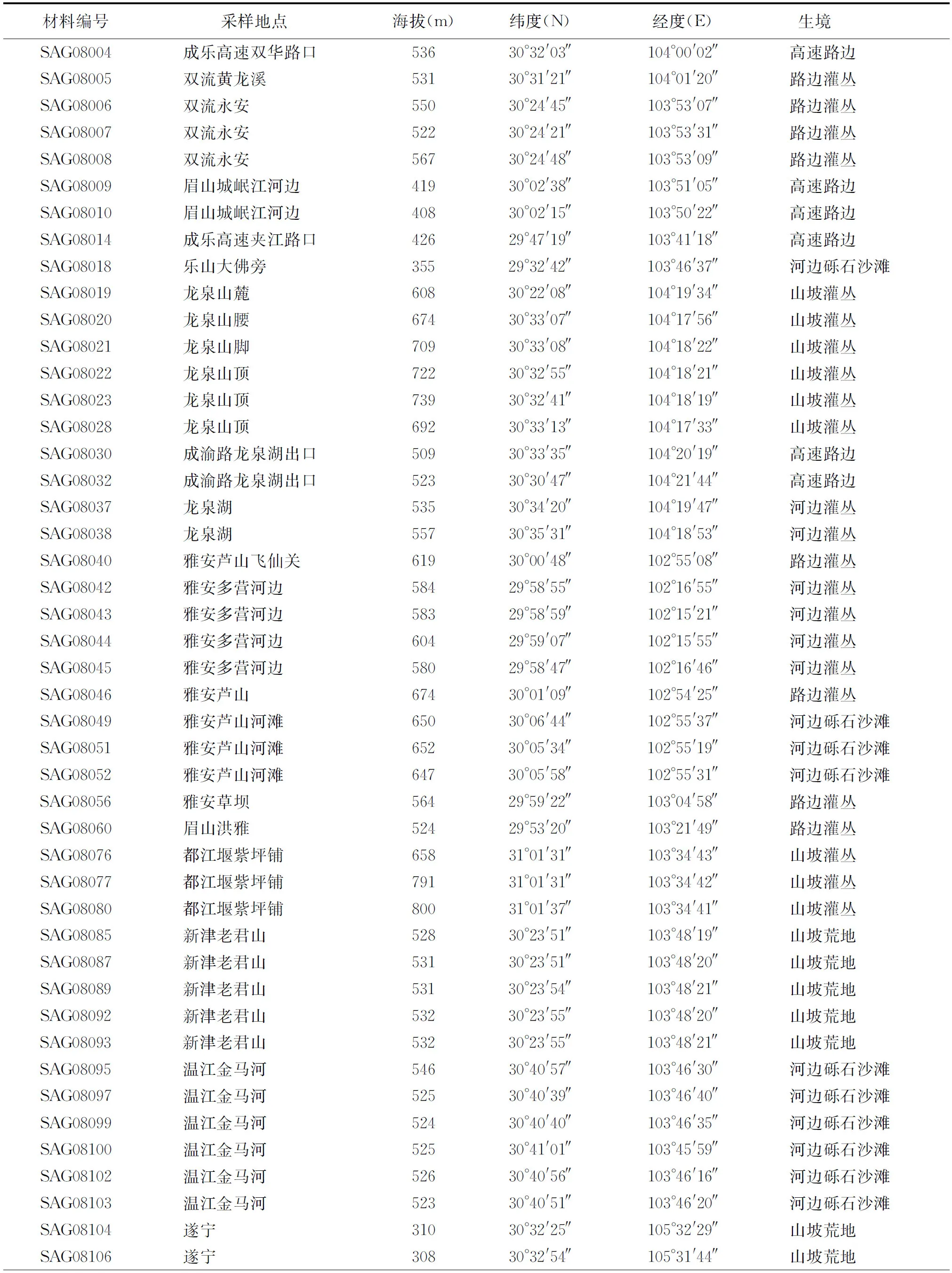
表1 60份斑茅種質資源的來源和生境
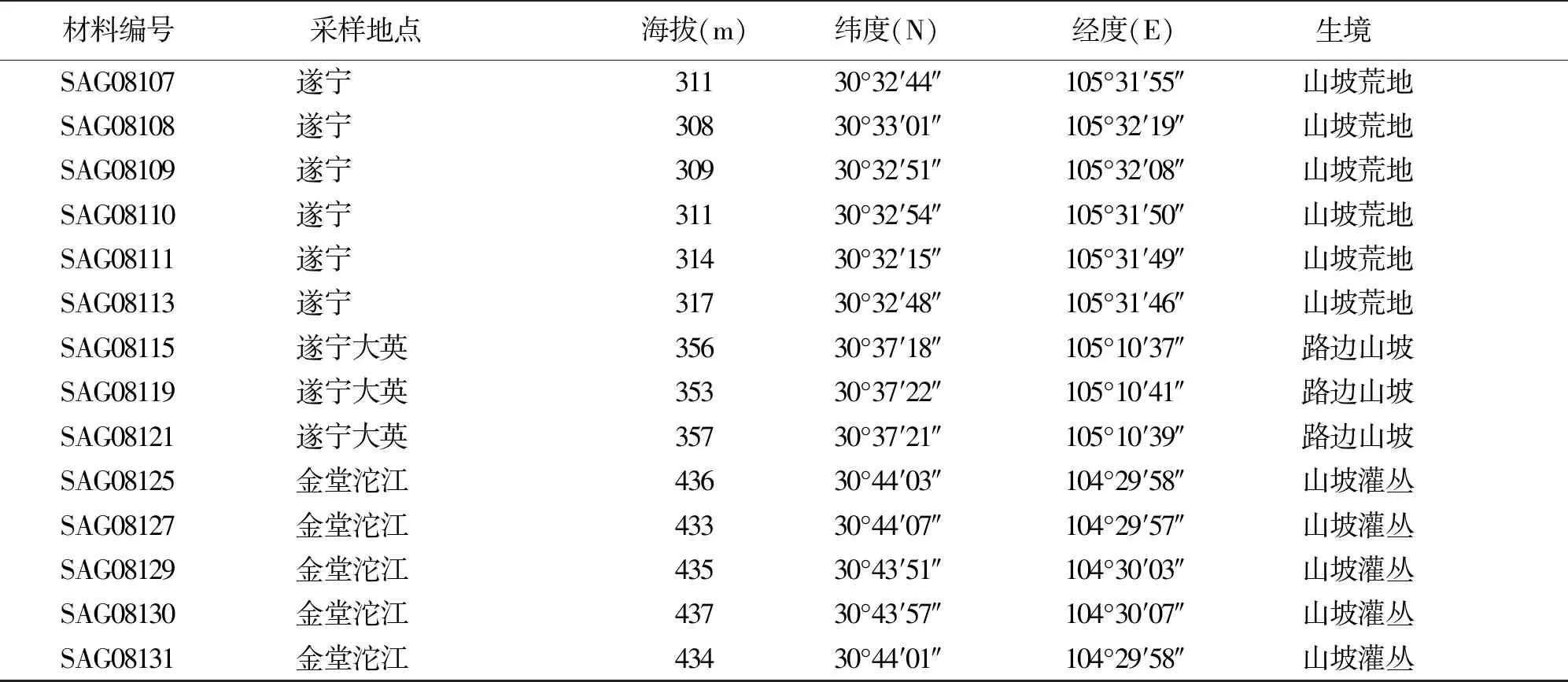
續(xù)表1
1.4.2穗部性狀 在蠟熟期每個指標選擇5~10個單株測定穗長、穗節(jié)數、穗中部節(jié)上的小穗數、小穗長,其中穗長、小穗長用卷尺測量。
1.5數據分析 利用Excel和SPSS軟件對材料的各形態(tài)指標數據進行描述性分析,包括變異范圍,平均數,變異系數、標準差等;對各形態(tài)指標進行方差分析,了解不同材料的表型性狀變異,通過相關分析確定各性狀之間的相互關系。利用NTSYS軟件基于各形態(tài)指標的平均數計算供試材料間的歐氏距離,并進行UPGMA聚類分析,了解各種質之間的親緣關系,將各材料劃分為在某些形態(tài)學特性上差異顯著的幾個大類群。
2 結果
2.1斑茅形態(tài)性狀變異分析 對60份斑茅種質資源11個形態(tài)學特征的統(tǒng)計表明,不同材料之間存在很大差異,表現出明顯的形態(tài)多樣性(表2)。變異幅度最大的是莖粗,變異范圍為8.86~37.36 mm,變異系數高達49.66%;其次是旗葉長,變異系數為45.10%;倒二葉寬和穗中部節(jié)上的小穗長的變異系數也很高,分別為44.20%和42.70%;而變異幅度最小的是穗節(jié)數,變異系數為16.72%。由此可見,斑茅種質營養(yǎng)器官的變異較大,其影響生產性能的特征具有很強的可塑性,所以可以結合斑茅的農藝性狀和生產性能選擇植株高大的。而斑茅的生殖器官除穗中部節(jié)上的小穗長外的指標都變異較小,可以采用無性繁殖的方法選擇優(yōu)良植物。
2.2斑茅形態(tài)學特征的相關性分析 生物變異是與環(huán)境條件相結合的,而且某些性狀之間的變異是相互聯系的。為了分析斑茅形態(tài)特征之間的相互關系,對不同來源的斑茅材料的11個指標進行了相關性分析。
2.2.1營養(yǎng)器官的相關性 營養(yǎng)器官各性狀間,旗葉長與旗葉寬、倒二葉長和倒二葉寬呈極顯著正相關,相關系數分別為0.575、0.614和0.338(表3);旗葉寬與倒二葉長和寬呈極顯著正相關,相關系數分別為0.400和0.472;倒二葉長與倒二葉寬呈極顯著正相關,相關系數為0.464。倒二葉長與株高呈顯著正相關,相關系數為0.100;倒二葉寬與株高呈顯著正相關,相關系數為0.113。莖粗與株高和莖節(jié)數呈極顯著的正相關,相關系數分別為0.216和0.139。株高與莖節(jié)數呈極顯著正相關,相關系數為0.547。由此可見,旗葉的長寬和倒二葉的長寬是彼此正相關的,株高、莖粗、莖節(jié)數也是彼此正相關,倒二葉的長寬與株高呈正相關,隨著株高增加,莖節(jié)數增加,莖粗也在增加。
2.2.2生殖器官的相關性 各生殖器官性狀之間,穗長與穗節(jié)數、穗中部節(jié)小穗數、穗中部小穗長之間呈顯著正相關,相關系數分別為0.383、0.122和0.092。穗節(jié)數與穗中部小穗數呈顯著正相關,相關系數為0.091。穗中部小穗數和穗中部小穗長呈極顯著正相關,相關系數為0.173。穗中部小穗數越多,穗中部小穗就越長。

表2 供試斑茅種質資源形態(tài)學特性及其變異
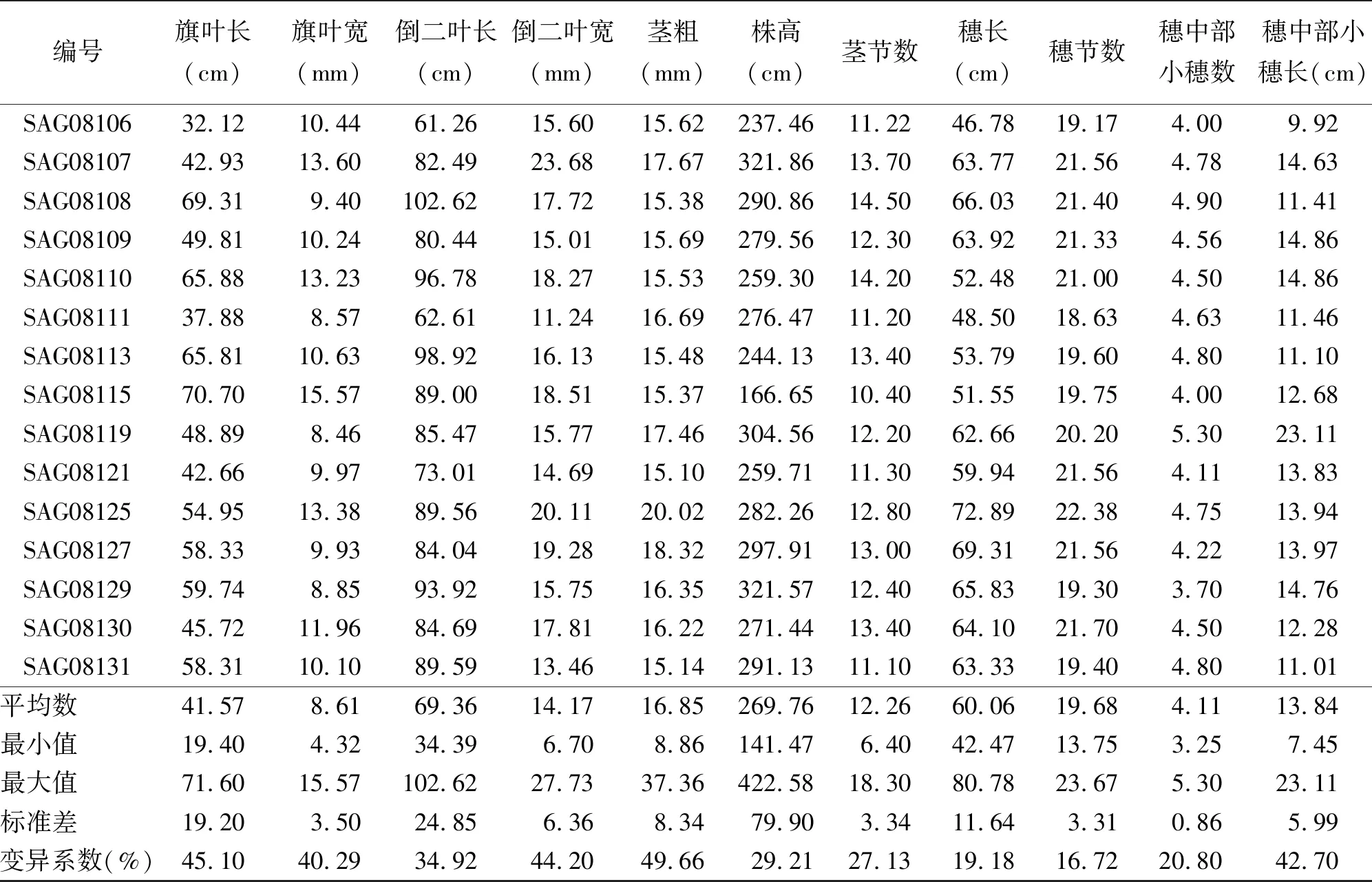
續(xù)表2
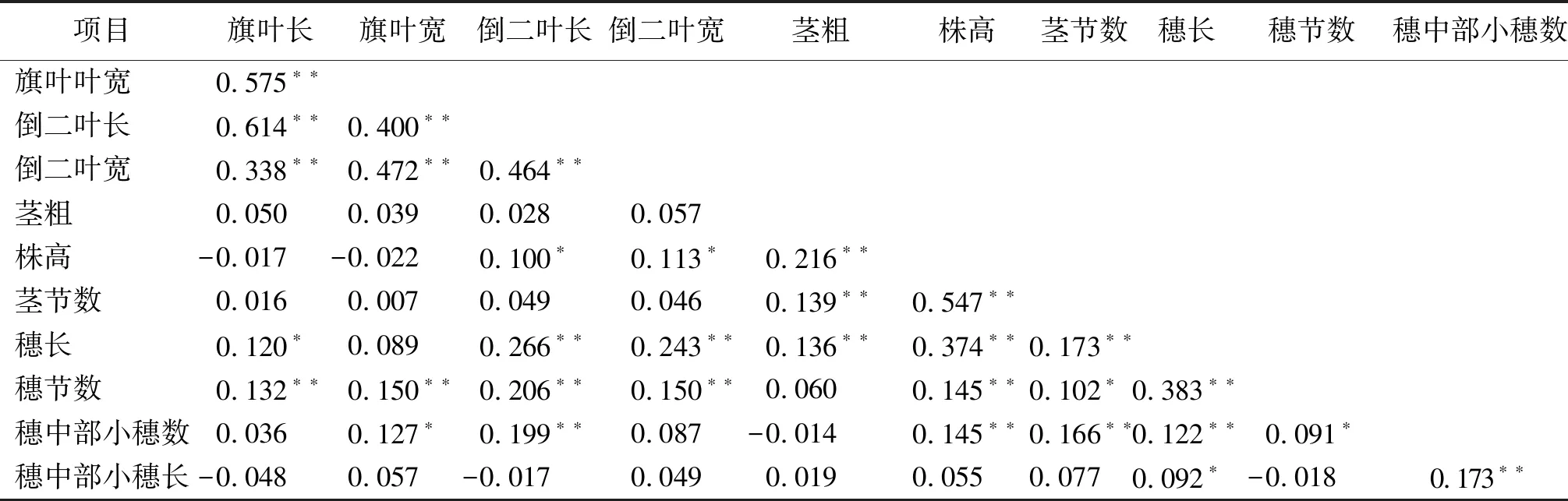
表3 斑茅種質資源形態(tài)學性狀間的相關關系
結合營養(yǎng)器官與生殖器官的相關關系可見,營養(yǎng)器官呈現為植株高大,莖較粗,莖節(jié)數多,旗葉葉片較寬的斑茅,穗長也長,穗中部小穗數及小穗長也增加。所以通過斑茅形態(tài)特征的早期選擇可以獲得植株高大的斑茅,也可以獲得種子生產潛力較好的種源來研究斑茅種子的特性。
2.3斑茅形態(tài)學特征的聚類分析 對供試的60份斑茅材料依據11個植物學形態(tài)特性進行聚類分析,可將其劃分為3個形態(tài)學類型(圖1)。第Ⅰ類包括SAG08004、SAG08006、SAG08007、SAG08008、SAG08014、SAG08042、SAG08043、SAG08044、SAG08049、SAG08052、SAG08076、SAG08078、SAG08080、SAG08085、SAG08089、SAG08097、SAG08099、SAG08100、SAG08102、SAG08107、SAG08108、SAG08109、SAG08111、SAG08119、SAG08125、SAG08127、SAG08129、SAG08130和SAG08131。這一類植株比較高大,莖稈較粗,穗長也較長,葉子長而寬,莖節(jié)和穗節(jié)數也較多。第Ⅰ類材料的幾乎所有性狀都是參試材料中表現最好的,各性狀的平均值高于所有材料的總體平均值,說明該類材料作為能源植物具有較大的開發(fā)潛力(表4)。
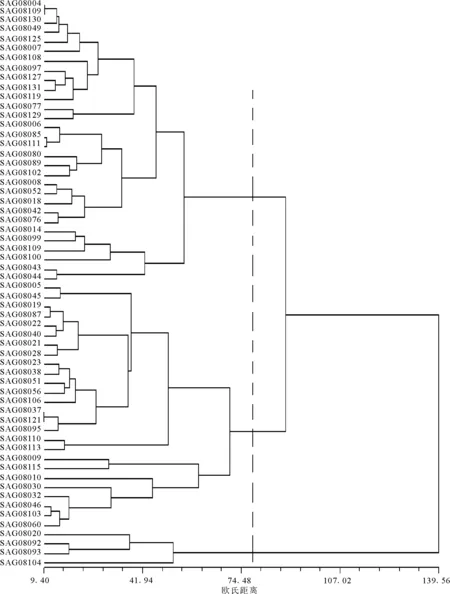
圖1 斑茅種質資源形態(tài)學性狀聚類樹系
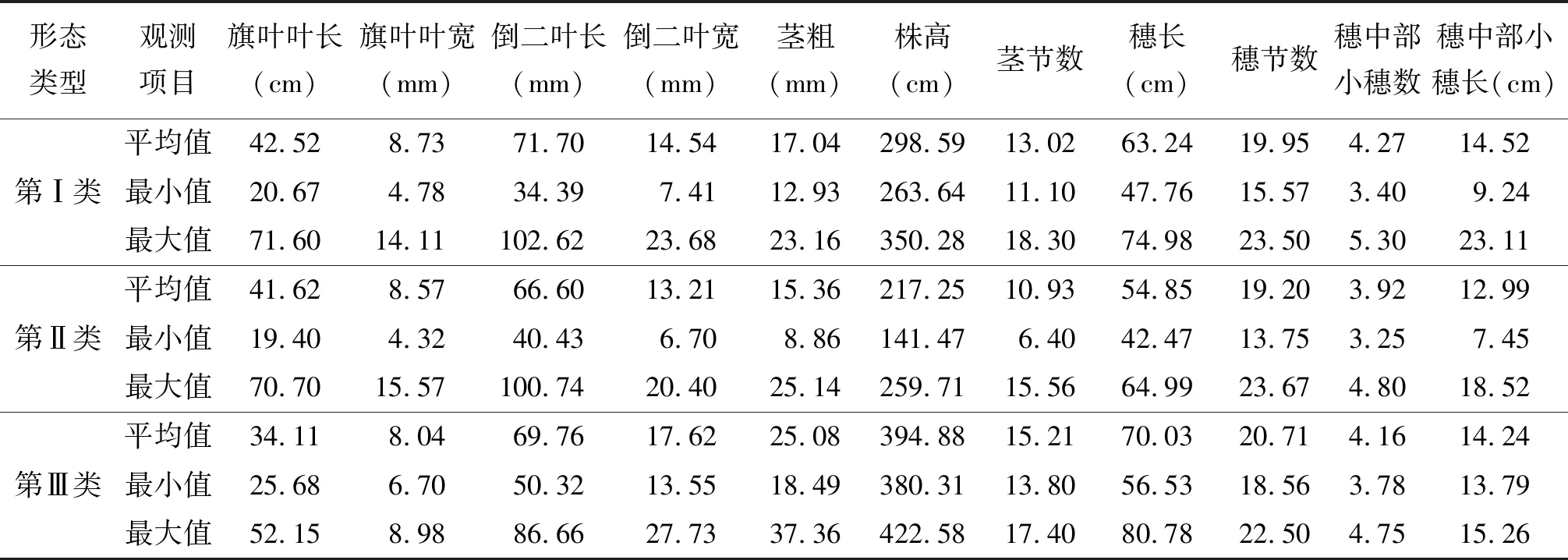
表4 斑茅三大類群的形態(tài)特征
第Ⅱ類材料包括SAG08005、SAG08009、SAG08010、SAG08019、SAG08021、SAG08022、SAG08023、SAG08028、SAG08030、SAG08032、SAG08037、SAG08028、SAG08040、SAG08045、SAG08046、SAG08051、SAG08056、SAG08060、SAG08087、SAG08095、SAG08103、SAG08106、SAG08110、SAG08113、SAG08115和SAG08121。第Ⅱ類材料在形態(tài)學上表現的特征為:植株矮小,莖稈較細,穗長較短,葉短而窄。第Ⅱ類材料的幾乎所有性狀都是參試材料中表現較差的,各性狀的平均值略低于所有材料的總體平均值,此類材料可不作為能源植物開發(fā)。
第Ⅲ類材料為SAG08020、SAG08092、SAG08093和SAG08104。此類的4個材料各個形態(tài)指標表現最差。但是這4個材料的株高、莖粗和莖節(jié)數都很高,可用來改良第Ⅰ類材料植株的性狀。
以上聚類分析結果可見,形態(tài)相似的斑茅首先聚在一起,聚類結果與材料的來源無明顯一致性,但是具有一定的相關性。
3 討論與結論
3.1不同種質資源的形態(tài)特征存在廣泛的變異 形態(tài)性狀具有穩(wěn)定性和變異性,受本身的遺傳組成和所處環(huán)境兩方面的影響,盡管形態(tài)變異具有一定的遺傳基礎,但是環(huán)境壓力在導致形態(tài)變異中也起著重要的作用[13-15]。植物很難處于生長發(fā)育最適宜的環(huán)境條件,無論是氣候變化還是生長發(fā)育過程的群落條件,總是要產生或大或小的差異[16-17]。本研究表明,不同種源的野生斑茅存在廣泛的形態(tài)差異,11個指標變異程度大小順序為:莖粗>旗葉長>倒二葉寬>穗中部小穗長>旗葉寬>倒二葉長>株高>莖節(jié)數>穗中部小穗數>穗長>穗節(jié)數。由此可見,穗部特征除了穗中部小穗長變異程度較大外,其他特征變異程度均較小;營養(yǎng)器官的變異都很大,可塑性較強,有很多作為能源植物的優(yōu)良性狀,所以應重視其作為能源植物的開發(fā)。
3.2各形態(tài)性狀之間顯著相關 在營養(yǎng)器官各性狀之間,隨著株高增加,莖節(jié)數也在增加,葉子長而寬,莖粗也隨著增粗,反之亦然。4個生殖器官指標之間呈顯著的相關性,穗長越長,穗節(jié)數越多,穗中部小穗數就越多。在營養(yǎng)器官與生殖器官性狀間也存在顯著關系,株高越高,莖稈越粗,莖節(jié)數越多,葉片長而寬,穗長就越長,小穗也越多且長。
3.3聚類結果與形態(tài)相似性有關 源自不同地區(qū)斑茅材料,形態(tài)相似的首先聚在一起,聚類結果與材料的來源無明顯一致性,但是具有一定的相關性。本研究供試材料分為3類,第Ⅰ類材料較多,包括了來自雙流、樂山、雅安、溫江、都江堰、遂寧以及金堂的,其中遂寧、溫江以及金堂的材料較多,包括SAG08097、SAG08099、SAG08100、SAG08102、SAG08107、SAG08108、SAG08109、SAG08111、SAG08119、SAG08125、SAG08127、SAG08129、SAG08130和SAG08131。該類材料所有性狀都是參試材料中表現最好的,各性狀的平均值高于所有材料的總體平均值,所以應該結合第Ⅲ類材料作為能源植物開發(fā)的重點。
[1]中國科學院中國植物志編委會.中國植物志(第十卷第二分冊)[M].北京:中國農業(yè)出版社,1997:40-45.
[2]南京大學生物系,中科院植物所.中國主要植物圖說(禾本科)[M].北京:科學出版社,1965:758.
[3]四川植物志編輯委員會.四川植物志(第十二卷)[M].成都:民族出版社,1998:337.
[4]廖兆周,勞方業(yè),周耀輝,等.具有斑茅種質的耐旱甘蔗品系的選育[J].作物學報,2002,28(6):841-846.
[5]李奇?zhèn)ィ饕?甘蔗×斑茅雜種及其雙親的農藝、工藝、植物學性狀和染色體觀察[J].云南農業(yè)大學學報,1995,10(1):29-34.
[6]余愛麗,張木清,陳如凱.ISSR分子標記在甘蔗及其近緣屬分類上的應用[J].福建農林大學學報(自然科學版),2002,31(4):484-489.
[7]王洪新,胡志昂,鐘敏,等.毛烏素沙地錦雞兒種群形態(tài)變異[J].生態(tài)學報,1994,14(4):366-370.
[8]葛頌,洪德元.泡沙參復合體(桔梗科)的物種生物學研究:Ⅰ.表型的可塑性[J].植物分類學報,1994,32(6):489-503.
[9]孟林,張國芳,高洪文.氮磷鉀施肥量對飼用菊苣生產性能的影響[J].草地學報,2003,11(4):325-328.
[10]葛頌,洪德元.泡沙參復合體(桔梗科)的物種生物學研究[J].植物分類學報,1995,33(5):433-443.
[11]葛頌,洪德元.瀕危物種裂葉沙參及其近緣廣布種泡沙參的遺傳多樣性研究[J].遺傳學報,1999,26(4):410-417.
[12]Kellogg E A.Variation and names in thePoasecundacomplex[J].Journal of Range Management,1997,38(6):516-521.
[13]嚴學兵,郭玉霞,周禾,等.青藏高原垂穗披堿草變異的地理因素分析[J].西北植物學報,2007,27(2):328-333.
[14]楊繼.植物種內變態(tài)的機制及其研究方法[J].武漢植物研究,1991,20(4):429-434.
[15]王金龍,高玉葆,趙念席,等.內蒙古中東部草原克氏針茅形態(tài)特征和RAPD遺傳分析的相關性分析[J].植物研究,2006,26(6):709-714.
[16]云錦鳳,王勇,徐春波,等.新麥草新品系生物特征及生產性能研究[J].中國草地學報,2006,28(5):1-7.
[17]楊允菲,李建東.東北羊草草原種群單穗數量性狀的生態(tài)可塑性[J].生態(tài)學報,2001,21(5):753-758.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
