沙打旺苗期對黃矮根腐病菌的抗性評價
2011-04-25 09:39:44俞斌華南志標李彥忠
草業科學 2011年7期
俞斌華,南志標,李彥忠
(蘭州大學草地農業科技學院,甘肅 蘭州 730020)
沙打旺(Astragalusadsurgens)是豆科黃芪屬多年生草本植物,屬于直根深根系植物,因其抵御風沙能力強而得到“沙打旺”的美譽[1]。沙打旺作為優質牧草,具有耐鹽堿、耐瘠薄、抗寒和抗旱等特性,20世紀70年代初,在我國黃土高原被大面積種植[2],到1986年底,在隴東黃土高原栽培面積近8.2萬hm2[3],現廣泛分布于我國東北、華北、西北等地[4]。在俄羅斯、日本、蒙古以及北美洲溫帶地區也廣泛分布[5]。
沙打旺雖為多年生牧草,但其草地利用年限較短,經濟利用期僅有2~3年,草地建植3年后,蓋度從100%降低到60%,5年后開始出現衰退現象[6-8]。真菌性病害是造成沙打旺產量、品質降低和草地利用年限縮短的主要原因[9-12]。由沙打旺埃里磚格孢(Embellisiaastragalisp. nov. Li & Nan)引起的黃矮根腐病是沙打旺植株上新發現的最嚴重的病害之一,其不僅使沙打旺草地建植不良,而且使沙打旺草地的持久性縮短,嚴重影響沙打旺草地的生產力、利用壽命、牧草品質和種子產量[13-15]。該病菌可寄生于植物全株,在葉柄和莖稈中的分離率顯著大于葉片和根部,引起植株系統發病,典型癥狀為葉斑、葉片黃化、莖稈矮化、莖稈變色、根頸和根腐爛,只在死亡植株表面產生分生孢子。病菌在種皮內、病株的莖基部、根頸、根中以及在死亡植株表面越冬,通過種子帶菌遠距傳播,死亡病株上的分生孢子通過氣流和雨水飛濺傳播,在春秋兩季發生侵染,病株帶菌引起植株連年發病。土壤水分越高、植株越稠密,危害越重[13-14,16]。
用于控制植物病害的方法主要有:利用抗病品種、物理防治、生物防除及殺菌劑處理。使用殺菌劑和生物防除方法成本較高,尤其是使用大量化學性殺菌劑,還會污染環境,威脅生物安全。與之相比,利用抗病品種是控制植物病害最經濟、最安全、最有效的方法[17-19]。
然而,目前關于沙打旺抗黃矮根腐病的抗病品種篩選,國內外鮮見報道。因此,評價沙打旺對埃里磚格孢的抗病性對于沙打旺黃矮根腐病的防治具有重要的意義。本研究評價我國的10份沙打旺材料在幼苗階段對黃矮根腐病的抗病性,是溫室和田間抗病性評價的重要基礎工作,以期為通過利用抗病品種防治沙打旺黃矮根腐病提供理論依據。
1 材料與方法
1.1沙打旺種子 本試驗收集了中國沙打旺主要種植區的10份材料,分為育成品種、地方品種和野生材料(表1)。種子保存在蘭州大學草地農業科技學院種子庫中(4 ℃,黑暗),種子發芽率依照國家種子檢驗規程[20]操作。
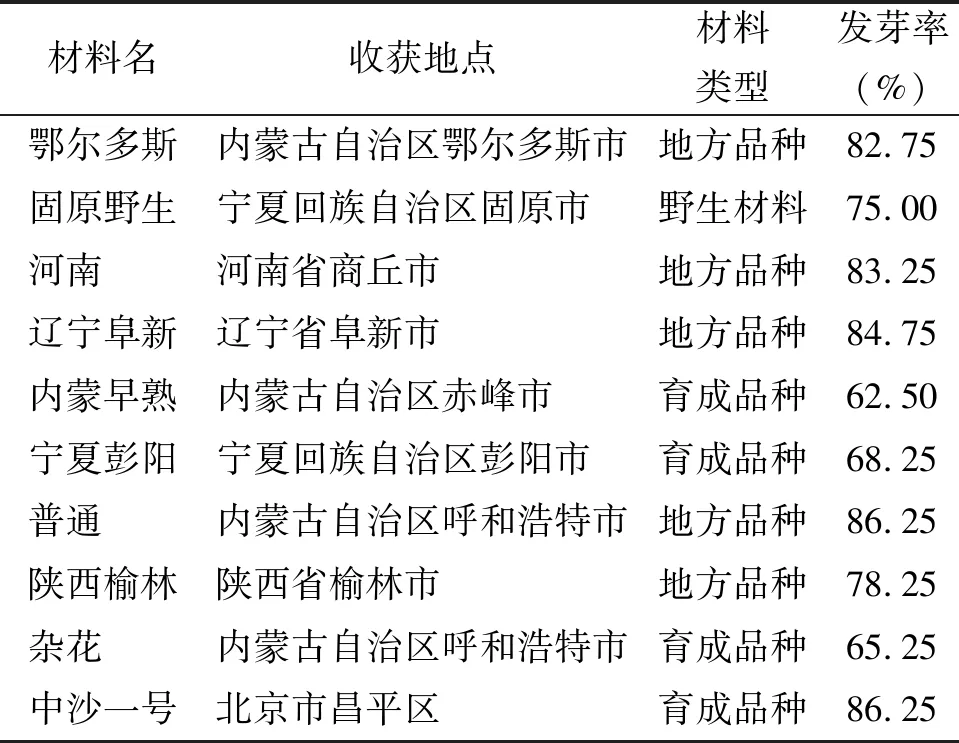
表1 沙打旺種子材料
1.2沙打旺埃里磚格孢 本試驗接種病原真菌為沙打旺埃里磚格孢模式種MHLZU-HX0401,分離自甘肅省環縣(37.12° N、106.82° E,海拔1 650 m)發病沙打旺植株莖部,并于22 ℃恒溫培養箱(Sanyo Incubator,Japan)中,WHDA(wheat hay decoction agar)培養基上密封培養3個月。
1.3接種方法 將沙打旺種子分別經過1%次氯酸鈉溶液處理10 min和75%酒精溶液處理3 min后,用無菌水沖洗3次,置于無菌濾紙上晾干[21]。然后用滅菌鑷子將種子均勻擺放在培養皿中埃里磚格孢菌落邊緣(接種處理),每皿10粒,每5皿一個重復,共5個重復,隨機擺放在22 ℃恒溫培養箱中黑暗培養。未接種處理為將表面消毒的種子放置于不接種的WHDA空白培養基表面培養。
1.4數據測量 按照國家種子檢驗規程[20],每天統計種子的發芽率,其中第3天的發芽率定義為種子的發芽勢[22]。
于第15天用直尺測量幼苗苗長、根長,并將幼苗以培養皿為單位置于80 ℃烘箱中烘36 h后[21],稱量幼苗干質量(Mettler Toledo,AL104),然后計算出接菌苗比未接菌苗干質量的降低比例。
1.5抗病性評價 每3 d隨機選取4皿接種幼苗,于顯微鏡(Leica DM 2500,200×)下測量菌絲體在幼苗表面上的擴展長度。然后將其截成數段(視幼苗根莖長度確定段數,每段0.5 cm),于0.05%的苯胺藍染液中沸煮15~30 s,用蒸餾水沖洗后在顯微鏡下(400×),測量幼苗根、莖中菌絲的分布長度和深度[23]。
于第15天計算發病率、死亡率,并依據變色與壞死部分的總長度占整個幼苗長度的比例評價癥狀強度(symptom intensity,SI):0=無變色與壞死,1=1.0%~10.0%,2=10.1%~30.0%,3=30.1%~50.0%,4≥50.1%,5=幼苗死亡[13]。然后用公式[24]計算出病情嚴重度(disease severity index,DSI)。
最后將所有材料依據病情嚴重度分類:免疫=0,抗病=0.1%~20.0%,中抗=20.1%~40.0%,中感=40.1%~60.0%,感病>60.0%[25-26]。
1.6統計分析 所有數據用Microsoft Excel 2003作圖,并用SPSS 13.0分析。
2 結果
2.1發芽率和發芽勢 所有材料接種沙打旺埃里磚格孢后的最初階段,發芽速度都有所增加,發芽勢均顯著高于未接種種子(P<0.05)(表2)。之后未接種種子的發芽率上升較快而接種種子的發芽率上升緩慢。接種后15 d時,所有沙打旺材料的發芽率均高于70%,其中8份材料的沙打旺種子在埃里磚格孢處理中發芽率均小于未接種種子,但差異不顯著(P>0.05)。埃里磚格孢對雜花和寧夏彭陽沙打旺種子的發芽率未產生影響(表2)。
2.2苗長和根長 與未接種幼苗相比,所有接種幼苗的苗長均有所降低,其中鄂爾多斯、河南、遼寧阜新、內蒙早熟、寧夏彭陽5個栽培品種以及固原野生沙打旺苗長的降幅均高于20%;而普通、陜西榆林、雜花和中沙一號苗長的降幅也接近10%,并且前6份材料與后4份材料苗長降幅差異顯著(P<0.05)(表3)。與未接種對照相比,埃里磚格孢顯著增加了所有沙打旺材料幼苗的根長(P<0.05),增幅均高于90%。其中鄂爾多斯、寧夏彭陽根長的增幅顯著高于河南和遼寧阜新根長的增幅(P<0.05),其余材料介于其間(表3)。
2.3幼苗干質量 埃里磚格孢顯著降低了10份沙打旺材料幼苗的干質量(P<0.05),其中鄂爾多斯、固原野生、河南、遼寧阜新、內蒙早熟和寧夏彭陽的幼苗干質量降幅均高于或接近40%,而普通、陜西榆林、雜花和中沙一號幼苗干質量的降幅均低于30%,并且前6份材料幼苗干質量的降幅顯著高于后4份材料的降幅(P<0.05)(表3)。

表2 埃里磚格孢對沙打旺種子發芽的影響 %
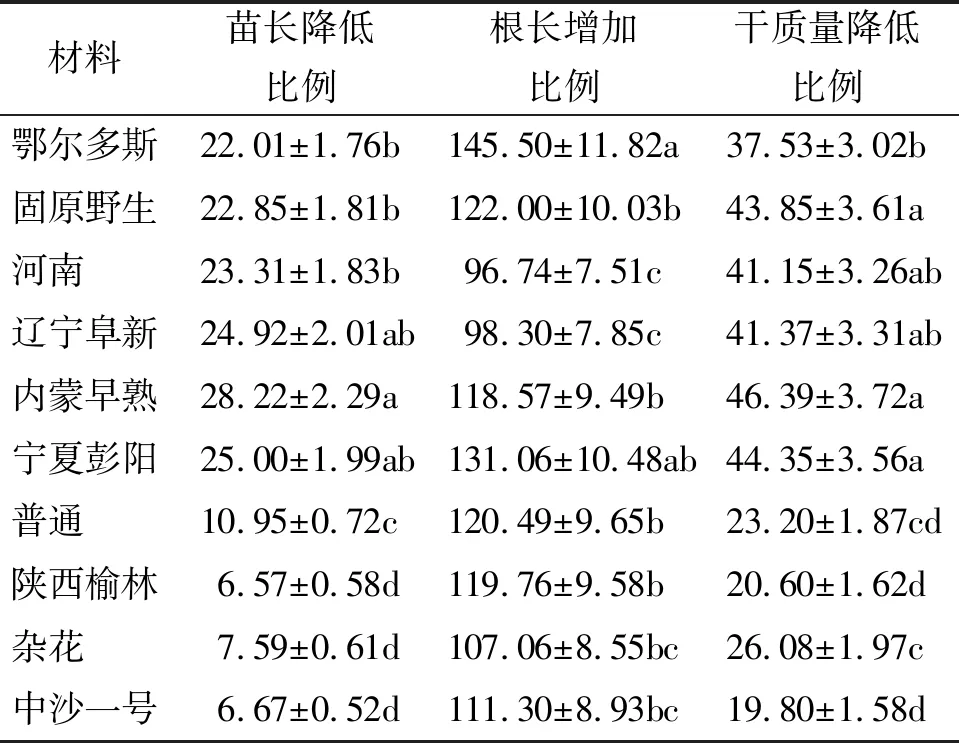
表3 埃里磚格孢對沙打旺幼苗苗長、根長及干質量的影響 %
2.4病原菌擴展水平 在10份沙打旺材料的接種幼苗上,埃里磚格孢菌絲都呈擴展生長的趨勢。5次測量,各材料幼苗上菌絲擴展長度均差異顯著(P<0.05)。菌絲擴展長度由大到小依次為:內蒙早熟、寧夏彭陽、固原野生、遼寧阜新、河南、鄂爾多斯、普通、雜花、陜西榆林、中沙一號(表4)。其中,埃里磚格孢菌絲在固原野生、遼寧阜新、內蒙早熟、寧夏彭陽沙打旺上的擴展長度顯著大于在鄂爾多斯、普通、陜西榆林、雜花、中沙一號上的擴展長度(P<0.05)。不同測量時間,同一材料幼苗上的菌絲擴展長度也差異顯著(P<0.05)(表4)。
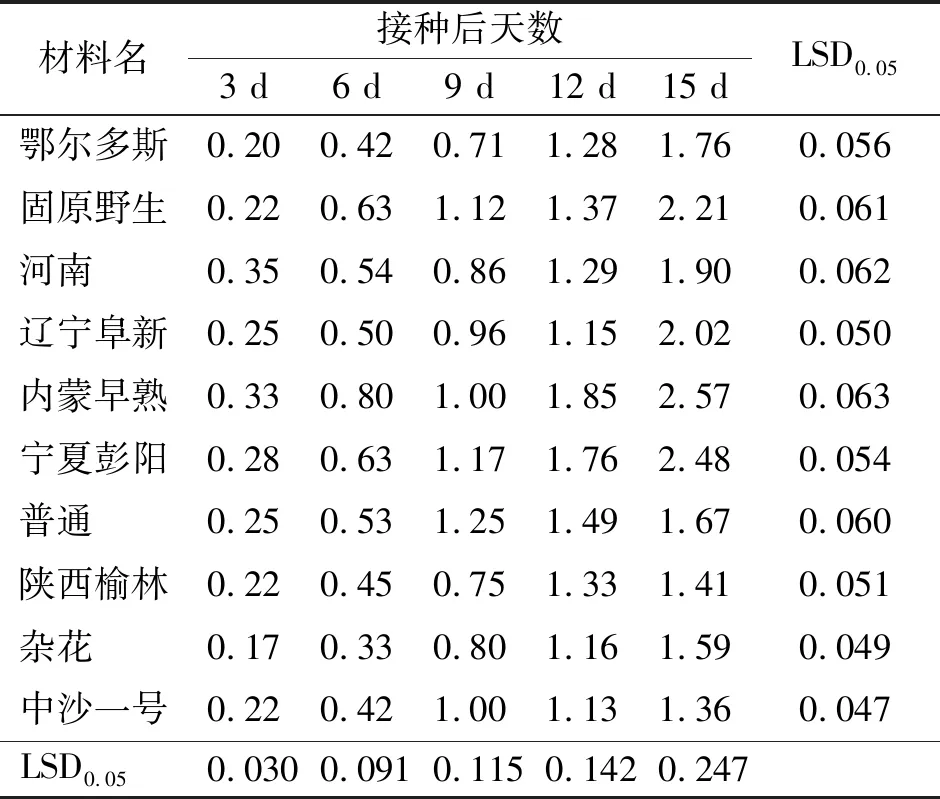
表4 接種沙打旺埃里磚格孢后幼苗中菌絲的擴展長度 cm
2.5病原菌侵入水平 在沙打旺幼苗中,埃里磚格孢菌絲也向幼苗內部入侵。接種后3 d時所有材料均有超過58%的幼苗表面分布有埃里磚格孢菌絲;接種后6 d時10個沙打旺材料中均有超過3%幼苗的皮層中檢測到埃里磚格孢菌絲;接種后15 d時埃里磚格孢菌絲侵入沙打旺各材料幼苗髓部,侵染率超過5%(表5)。其中,侵入速度較快的有鄂爾多斯、固原野生、內蒙早熟和寧夏彭陽沙打旺,侵入速度較慢的有普通、陜西榆林、雜花和中沙一號沙打旺。接種后15 d時在固原野生、內蒙早熟和寧夏彭陽沙打旺的部分幼苗上觀察到菌絲已經于根莖處產孢(表5)。
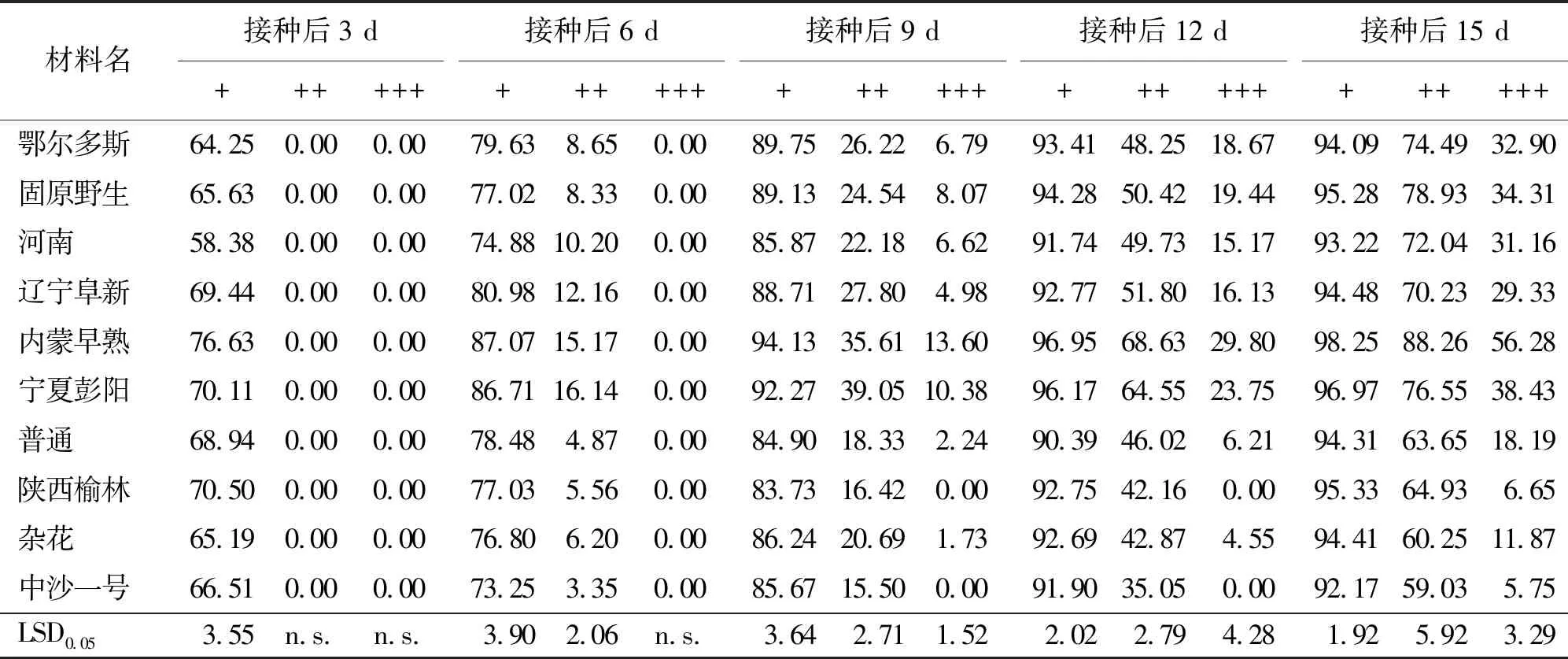
表5 沙打旺幼苗在埃里磚格孢菌絲侵入不同水平下的侵染率 %
2.6抗病評價 所有材料的幼苗在接種后發病率、死亡率、癥狀強度和病情嚴重度均差異明顯。內蒙早熟和寧夏彭陽的發病率、死亡率、癥狀強度和病情嚴重度均顯著高于其他材料(P<0.05),而陜西榆林和中沙一號的各項抗病相關指標則具有較低的值(表6)。通過分析以上指標可以得到如下結果:在10個沙打旺材料中,無免疫材料;抗病材料為陜西榆林和中沙一號;中抗材料為普通和雜花;中感材料為鄂爾多斯、固原野生、河南和遼寧阜新;感病材料為內蒙早熟和寧夏彭陽(表6)。
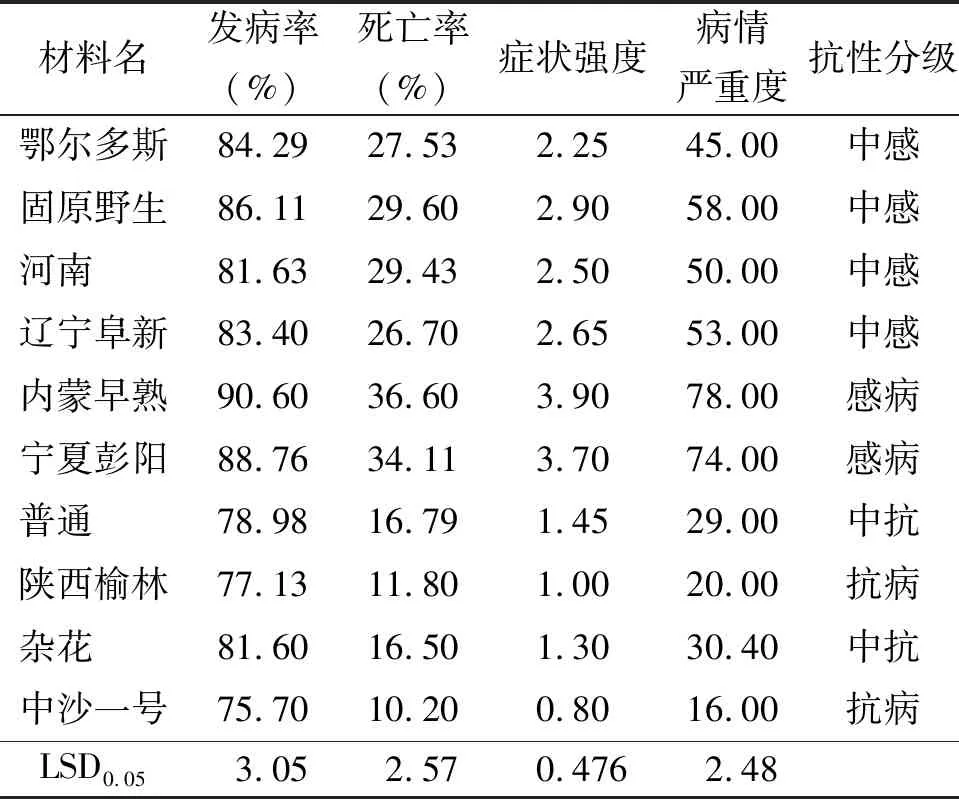
表6 發病率、死亡率、癥狀強度、病情嚴重度和抗性分級
3 討論
本研究在室內評價了10份沙打旺材料對黃矮根腐病的抗病性。根據埃里磚格孢對沙打旺種子發芽率、發芽勢、苗長、根長、苗干質量、菌絲在幼苗中的擴展和侵入水平等指標的綜合分析,得出在幼苗階段抗病性較強的材料為陜西榆林和中沙一號,抗病性較弱的材料為內蒙早熟和寧夏彭陽,其余材料介于其間。
埃里磚格孢未顯著降低參試沙打旺材料的發芽率,但在接種初期短時間內加速了種子的萌發,提高了存活率,與Li和Nan[15]的研究結果一致。埃里磚格孢顯著提高了10份沙打旺材料幼苗的發芽勢,這可能與病原菌影響了寄主的激素平衡或與病原菌產生的次生代謝物有關[27]。根苗長的變化是植株感病后的農藝性狀表現,也是不同沙打旺材料抗病性評價的重要依據[28]。本研究發現,病原菌顯著降低了其中6份沙打旺材料的苗長,與Li和Nan[15]的研究結果一致,但所有材料的幼苗根長顯著增加。健康幼苗的根較短,粗壯,但感病幼苗的根顯著增長、變細,有背向菌落中心生長的趨勢,并且與不同材料間抗病水平差異的趨勢不同。這可能與病原菌受到侵染后,植株中赤霉素總量減少,脫落酸濃度偏高,或其他激素平衡受到擾動有關,而與植物抗病水平無關[29],具體原因有待進一步研究。同時,埃里磚格孢侵染導致所有材料幼苗干質量顯著下降,是發芽率降低、死亡率上升、株高下降以及病情指數上升導致的發病與壞死的綜合表現。
對病原菌菌絲擴展與侵入的觀察統計發現,本研究中陜西榆林和中沙一號對沙打旺埃里磚格孢的抗性較強,菌絲在這兩個材料的幼苗表面擴展較慢,菌絲侵入時,在不同水平下幼苗侵染率也低于其他材料。與之相反,內蒙早熟和寧夏彭陽在這兩個指標中表現較差。這與寄主的抗侵入和抗擴展機制有關。當病原菌侵染時,寄主表皮細胞壁通常膨大,有時形成類膠狀沉淀層,而在抗病材料根部的壁旁區,會積累大量的多糖,并伴隨萜類和酚類化合物的合成,導致細胞壁增厚[29]。當病原菌侵入后,植株維管組織的細胞會表現壞死現象,壞死細胞可能導致寄主積累膠狀物質、胼胝質、木質素、木栓質和酚類物質,這些物質可限制病原菌對維管組織的進一步侵染[31-32],但具體機理尚需深入研究。
發病率、死亡率以及病情嚴重度是評價植物抗真菌病害的常用指標。本研究中內蒙早熟和寧夏彭陽的發病率、死亡率和病情嚴重度都顯著高于其余材料,結合其他數據可以得出這兩份材料屬于感病材料。說明這些指標直接反映了幼苗對病原菌的抗性水平,并能夠在溫室及田間條件下植物抗真菌病害評價中廣泛應用,如羽扇豆(Lupinusalbus)[33]、印度黃檀(Dalbergiasissoo)[25]及冬小麥(Triticumaestivum)[26]的抗病研究中都有廣泛使用。
本研究中,種子在萌發時就與埃里磚格孢菌落接觸,加速了侵染過程并提高了侵染強度,突出植株發病癥狀,使得研究結果真實可靠。本研究評價了10份沙打旺材料對埃里磚格孢的抗病性,初步篩選出抗病與感病材料,是今后溫室、田間抗病評價的重要基礎工作。另外,不同材料的幼苗抗病機理尚不明確,也是今后應予以重點研究的方向。
[1]蘇盛發.沙打旺[M].北京:農業出版社,1985.
[2]伊虎英,魚宏斌.輻射誘發超導早熟沙打旺的選育[J].草業科學,1989,6(3):18-21.
[3]甘肅畜牧廳.甘肅省牧草區劃[M].北京:中國農業出版社,1991.
[4]聶素梅,孫啟忠,閆志堅,等.影響沙打旺生長衰退因素試驗研究[J].內蒙古草業,2002(3):36-38.
[5]Frame J.Astragalusadsurgens[A].In:Frame J.Forage Legumes for Temperate Grasslands[M].Rome,ltaly and Enfield,NH,USA:FAO and Science Publishers,2005:51-54.
[6]孫啟忠,桂榮.科爾沁沙地沙打旺草地衰退規律與恢復技術研究[J].草地學報,2000,8(4):253-261.
[7]關秀琦,鄒厚遠,魯子瑜,等.黃土高原草地生產持續發展研究Ⅰ.沙打旺人工草地衰退后的草種更替[J].水土保持研究,1994(3):56-60.
[8]蘆滿濟,杜福成,楊志愛.冷溫半干旱黃土丘陵區荒坡地沙打旺系統生態效能的調查研究[J].草業科學,1994,11(2):48-51.
[9]孫啟忠,桂榮,那日蘇,等.赤峰地區不同生長年限沙打旺生產力的研究[J].中國草地,1999(5):29-34.
[10]趙明軒.慶陽地區沙打旺病害調查報告[J].草與畜雜志,1995(1):33-34.
[11]尹亞麗,南志標,李春杰,等.沙打旺根部入侵真菌的研究[J].草原與草坪,2006(1):45-48.
[12]南志標.沙打旺種帶真菌——環境,致病力及防治[J].草業學報,1998,7(1):12-18.
[13]Li Y Z,Nan Z B,Hou F J.The roles of anEmbellisiasp.causing yellow stunt and root rot ofAstragalusadsurgensand other fungi in the decline of legume pastures in northern China[J].Australasian Plant Pathology,2007,36:397-402.
[14]Li Y Z,Nan Z B.A new species ofEmbellisiacausing a disease of standing milk-vetch in China[J].Mycologia,2007,99(3):406-411.
[15]Li Y Z,Nan Z B.Symptoms and etiology of new disease,yellow stunt and root rot of standing milk-vetch in Northern China[J]. Mycopathologia,2007,163:327-334.
[16]Li Y Z,Nan Z B.Nutritional study onEmbellisiaastragali,a fungal pathogen of milk vetch (Astragalusadsurgens)[J].Antonie van Leeuwenhoek,2009,95:275-284.
[17]南志標.建立中國的牧草病害可持續管理體系[J].草業學報,2000,9(2):1-9.
[18]Chattopadhyay S,Ali K A,Doss S G,etal.Evaluation of mulberry germplasm for resistance to powdery mildew in the field and greenhouse[J].Journal of General Plant Pahtology,2010,7(6):87-93.
[19]王省芬,田海燕,馬峙英,等.黃萎病菌誘導下陸地棉抗病品種SSH文庫的構建[J].棉花學報,2008,2(1):3-8.
[20]王彥榮,孫建華,余玲,等.GB/T 2930.4-2001牧草種子檢驗規程發芽試驗[S].北京:中國標準出版社,2001.
[21]方中達.植病研究方法[M].北京:中國農業出版社,1998.
[22]鄭秀芳,李彩霞,周希豪,等.沙打旺種子萌發特性研究[J].種子,2007(6):43-45.
[23]李春杰,南志標.苜蓿種帶真菌及其致病性測定[J].草業學報,2000,9(1):27-36.
[24]Graham J H,Devine T E,Hanson C H.Occurrence and interaction of three species ofColletotrichumon alfalfa in the mid-Atlantic United States[J].Phytopathology,1976,66:538-541.
[25]Harsh N S K,Chandra S,Uniyal K.Screening resistance ofDalbergiasissooclones againstGanodermalucidumroot rot disease in field conditions[J].Forest Pathology,2011,41:221-226.
[26]Peterson G L.Reaction of selected winter wheat cultivars from Europe and United States to Karnal bunt[J].European Journal of Plant Pathology,2009,125:497-507.
[27]李彥忠,南志標.埃里磚格孢屬真菌的研究進展及展望[J].草業學報,2009,18(1):171-178.
[28]李彥忠.沙打旺(AstragalusadsurgensPall.)黃矮根腐病(Embellisiaastragalisp.nov.Li & Nan)的研究[D].蘭州:蘭州大學,2007.
[29]Matheussen A M,Morgan P W,Frederiksen R A.Implication of gibberellins in head smut (Sporisoriumreilianum) ofSorghumbicolor[J].Plant Physiology,1991,96:537-544.
[30]Daayf F,Nicole M,Boher B,etal.Early vascular defense reactions of cotton roots infected with a defoliating mutant strain ofVerticilliumdahliae[J].European Journal of Plant Pathology,1997,103(2):125-136.
[31]Bowers J H,Nameth S T,Riedel R M,etal.Infection and colonization of potato roots byVerticilliumdahliaeas affected byPratylenchuspenetransandP.crenatus[J].Phytopathology,1996,86(6):614-621.
[32]Linsell K J,Keiper F J,Forgan A,etal.New insights into the infection process ofRhynchosporiumsecalisin barley using GFP[J].Fungal Genetics and Biology,2011,48:124-131.
[33]Luckett D J,Cowley R B,Richards M F,etal.BreedingLupinusalbusfor resistance to the root pathogenPleiochaetasetosa[J].European Journal of Plant Pathology,2009,125:131-141.
