落葉松雜種與親本ISSR鑒別技術1)
2011-03-29 08:06:38張含國張磊
東北林業大學學報 2011年7期
張含國 張磊
(林木遺傳育種國家重點實驗室(東北林業大學),哈爾濱,150040)
朱航勇
(七臺河市園林管理局)
徐悅麗姚宇
(林木遺傳育種國家重點實驗室(東北林業大學))
落葉松(Larix)是一些生長快、木材用途廣、經濟價值高的樹種。長白落葉松(Larix Olgensis)、興安落葉松(L.gmelini)和日本落葉松(L.Kaempferi)分別具有不同的優良性狀:興安和長白落葉松具有良好的耐寒性,日本落葉松生長迅速且不易感染早期落葉病,其雜交種多數具有雙親的優良性狀。但不同的組合雜種優勢存在較大差異,遺傳增益差別很大[1-2],所以對雜種落葉松類別的鑒定十分必要。雖然從形態標記[3]、細胞學標記[4]和生化標記[5]等方面有關于落葉松種間的鑒別,但因局限性所致影響了其使用。由于從分子水平對家系間鑒別的研究很少,因而,對于主要靠有性繁殖的樹種來說,優良家系(雜種)的鑒別必不可少。自1994年加拿大蒙特利爾大學Zietkiewicz等[6]發展了ISSR標記以來,因其在分析和應用上比RFLP、AFLP和SSR更加簡便、快捷,而且又能比RAPD揭示出更多的多態性,現已在各種植物及林木的遺傳作圖、基因定位、遺傳多樣性等研究方面得到了廣泛應用,例如分析物種親緣關系[7-8]、遺傳作圖[9]或者與其它分子標記相結合進行相關分析[10]等。本研究采用ISSR分子標記技術研究日本落葉松、興安落葉松和長白落葉松及其子代的特異性,試圖找出適合的引物,通過不同雜種擴增條帶的差異,得到雜種和家系的鑒定方法,并根據遺傳距離對雜種家系進行聚類分析,希望能夠為不同雜種的準確鑒別提供新的途徑和手段。
1 材料與方法
材料來源——試驗材料來源于黑龍江省牡丹江市林口縣青山林場的落葉松種子園子代測定林。材料包括生長性狀優良的不同家系:興安落葉松×日本落葉松5個,興安落葉松×長白落葉松2個,日本落葉松×興安落葉松8個,日本落葉松×長白落葉松5個,共計20個家系,單株241個。材料具體名稱及數量見表1。采樣時采取當年生嫩葉(松針),置于冰上保存帶回,之后在冰箱中-40℃保存,以備提取樣本DNA。
植物基因組DNA提取——采用CTAB法提取植物總DNA,將用液氮研磨好的樣品加入CTAB中,65℃水浴后使用氯仿異戊醇抽提,無水乙醇清洗沉淀,最后將沉淀溶于適量TE中[11]。
反應體系和程序——落葉松ISSR反應體系為:DNTP(2.5 mmol/L)1.4 μL,Buffer2 μL,Mg2+(2.5 mmol/L)0.4 μL,引物2 μL,Tag酶0.2 μL,DNA模板2 μL。去離子水12 μL,總體積共20 μL。擴增程序為:94℃預變性3 min,然后進行35次循環,包括94℃變性30 s,56℃退火45 s,72℃延伸30 s,最后72℃總延伸7 min[12]。
擴增產物用含有溴化乙錠的1.5%瓊脂糖凝膠在1×TAE緩沖液中電泳分離,最后在紫外燈下用GENE GENIUS攝像系統照相記錄結果。
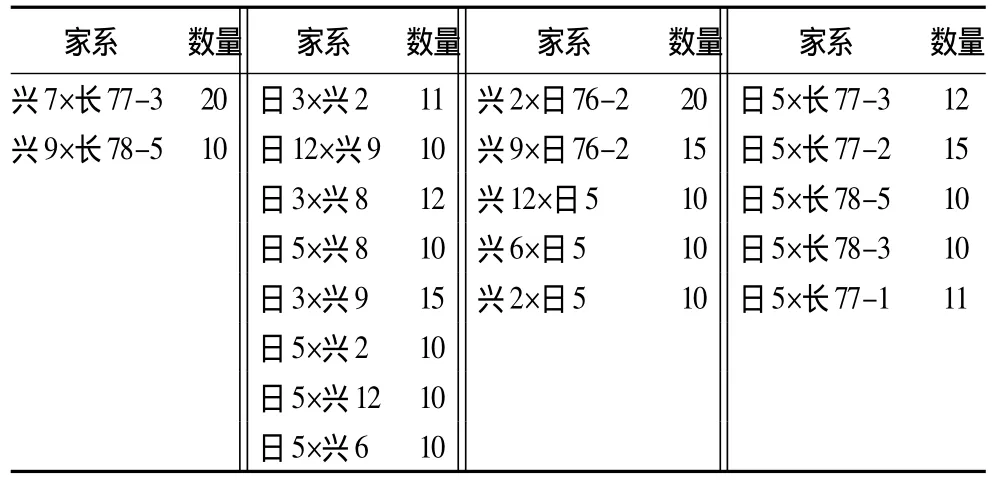
表1 家系名稱及子代個數
數據分析——對所有引物擴增的電泳結果進行統計分析,根據ISSR擴增產物相同電泳遷移位置條帶的有無,建立分子標記資料(多態性片段的數目、大小和信息量)的0、1數據。有帶記為1,無帶或模糊不清的條帶記為0。用所有引物重復擴增1~2次,選擇條帶清晰穩定、重復性好的擴增產物統計相對分子量大小,記錄結果。采用Popgen32軟件進行數據處理,計算家系間的遺傳距離,再以非加權兩兩對比法(Unweighted pair group methods of arithmetic(average),UPGMA)[13]建立聚類圖。
后期驗證——在取材地點隨機采集落葉松種及雜種的針葉,提取DNA,同時與實驗室已有的樣品一起用已篩選的引物進行擴增,然后記錄電泳圖片的數據,根據已有結果分析得出每個樣品的種類,最后與實際結果比較。
2 結果與分析
2.1 ISSR分子標記的鑒別
在落葉松種間及無性系間能夠用特異擴增條帶進行鑒別的基礎上[11],進一步進行雜種子代與親本、不同雜種組合間、雙親之一相同家系、正反交家系的鑒別。
引物編號及序列具體見表2。

表2 引物編號及堿基序列
2.1.1 不同雜種組合間鑒定
日本落葉松、興安落葉松和長白落葉松為父本和母本相互雜交,得到4種不同的雜種落葉松,分別為日×興、日×長、興×日、興×長。利用篩選好的ISSR引物對這4種不同的雜種落葉松進行擴增,對擴增結果分析發現:日×興和興×日的雜種落葉松很難區分,而日×長和興×長雜交的子代可以通過擴增條帶的特異性與其他雜種落葉松區別。
利用49個ISSR引物對不同雜種進行擴增,分析比較電泳圖片后發現:16號ISSR引物可以鑒定興安落葉松×長白落葉松。興×長的子代在16號引物擴增的條件下,在相對分子質量1 500 bp處擴增出特異譜帶,其它日×興、日×長和興×日的子代在這一位置均沒有出現譜帶(圖1)。
1 ~7.日本落葉松×興安落葉松;8~12.日本落葉松×長白落葉松;13~18.興安落葉松×日本落葉松;19~24.興安落葉松×長白落葉松;M.DL 2000 Marker。
19 號ISSR引物可以鑒定日本落葉松×長白落葉松。日×興、興×日和興×長的子代在相對分子質量900 bp處均有擴增條帶出現,而日×長白落葉松雜交的子代在19號引物的擴增下,在相同標準片斷處沒有任何擴增譜帶(圖2)。

圖1 16號引物對不同雜交組合的擴增結果
1 ~7.日本落葉松×興安落葉松;8~12.日本落葉松×長白落葉松;13~18.興安落葉松×日本落葉松;19~23.興安落葉松×長白落葉松;M.DL 2000 Marker。
2.1.2 雙親之一相同、家系不同的鑒定
試驗材料中有涉及雙親之一相同的不同家系。以日5為父本、母本不同的家系2個,以日3為母本、父本不同的家系3個。
日5為父本、母本分別為興6和興12的2個不同家系,在38號ISSR引物的擴增下共出現6條擴增譜帶。其中,在標準片段700 bp處,2個家系的子代擴增條帶出現的幾率有所不同。興6×日5的子代(圖3)出現擴增譜帶的幾率是88.89%,興12×日5的子代(圖4)在此處擴增出譜帶的幾率為10%。

圖2 19號引物對不同雜交組合的擴增結果

圖3 38號引物對興6、日5及其雜交子代的擴增結果

圖4 38號引物對興12、日5及其雜交子代的擴增結果
結果表明,在已知父本為日5、而母本不確定時,用38號引物對子代樣品進行擴增,根據700 bp擴增譜帶的有無可判斷家系的母本是哪個個體,如出現擴增譜帶,則樣品的母本為興6的可能性大于其母本為興12的可能性,其母本為興6的可能性為89.89%,其母本為興12的可能性為10.11%。如沒有擴增譜帶出現,則情況相反,其母本為興6的可能性為10.99%,其母本為興12的可能性為89.01%。
同樣,在引物ISSR36的擴增下,以日3為母本,父本分別為興2、興8和興9的3個不同家系的子代擴增出相對分子質量700 bp片段的幾率有所不同。具體統計結果如表3所示。

表3 日3為母本,父本不同的家系利用ISSR36擴增的檢測結果
2.1.3 正反交家系的鑒定
試驗材料中有由父母雙親正反交所產生的不同家系,在對這些家系的電泳圖片比較分析時發現,在不同ISSR引物的擴增下,有2對正反交組合可以進行區分。其中所涉及的4個家系每個家系各包括10個子代。
在用36號ISSR引物對由日5和興12正反交所產生的不同的家系進行擴增時發現,子代在400 bp處出現擴增片段,而父本和母本均沒有,其中興12×日5的子代出現此片段的特異率達到了100%,而日5×興12的子代出現此片段的特異率也達到90%(圖5、圖6)。并且,在相對分子質量700 bp處,興12×日5的子代在相同位置只有20%擴增出顏色非常淺的條帶,可以忽略不計(圖5),而日5×興12的子代有80%擴增出條帶(圖6)。這可以說明,當已知樣品為日5和興12雜交所得,但不確定父母本時,用36號引物對其進行擴增時,可以在700 bp處擴增出條帶的一定為日5×興12的子代;反之,如在此位置沒有擴增條帶出現時,則樣品為興12×日5的可能性為71.42%。

圖5 36號引物對興12、日5及其雜交子代的擴增結果

圖6 36號引物對日5、興12及其雜交子代的擴增結果
在用32號ISSR引物對由日5和興6正反交所產生的不同的家系進行擴增時發現,日5×興6的子代在相對分子質量350 bp處有擴增條帶出現,比率達到90%(圖7),而興6×日5的子代在此位置除1個子代外,其余均沒有擴增條帶出現(圖8)。結果表明:當用32號引物對已知為日5和興12雜交所得的子代進行擴增,但不確定父母雙親時,如果有350 bp擴增條帶出現,則樣品是日5×興6的可能性遠遠大于是興6×日5的子代,且其母本為日5的可能性為81.82%。

圖7 32號引物對日5、興6及其雜交子代的擴增結果

圖8 32號引物對興6、日5及其雜交子代的擴增結果
2.2 遺傳距離分析與雜種鑒別
以子代個體數相同的20個家系的遺傳距離進行計算,其遺傳距離的變幅及平均值分別為0.0161~0.4989和0.2642(表4、表5)。遺傳距離最小的是日5×長77-1和日5×長77-3家系,遺傳距離最大的是日3×興9和興9×日76-2家系。

表4 遺傳距離分析中家系代號
利用軟件繪制遺傳關系聚類圖。從家系間的遺傳關系聚類圖(圖9)可以看出,在0.4的遺傳距離上,可以將20個家系分為4類,分別是4種不同的雜交組合,即:日本落葉松×興安落葉松、日本落葉松×長白落葉松、興安落葉松×長白落葉松和興安落葉松×日本落葉松。遺傳距離的分類說明,不同落葉松雜種是可以進行區分的,與分子標記得到的結果基本吻合。

圖9 家系間遺傳關系聚類圖

表5 20個家系的遺傳一致度和遺傳距離
3 討論
目前,利用分子標記的方法對落葉松進行鑒定主要采取RAPD等方法,其中大多為種間的鑒定。主要研究有:管玉霞利用落葉松的部分葉綠體基因、線粒體基因和核糖體ITS序列,獲得了rbcL基因上的一個SNP標記,并結合線粒體特異片斷f-13、B-11可以對西部、美洲、日本和興安、長白落葉松做出準確快速的鑒別[14];曲麗娜等[15]用RAPD和ISSR分子標記對興安落葉松、長白落葉松及日本落葉松進行了不同物種間的鑒定。對于其它針葉樹種,雖然有人對無性系和雜交種進行過鑒別,但所用引物數量和所分析的無性系數量都較少[16]。張磊等[11]對長白落葉松、興安落葉松和日本落葉松的種間、無性系進行了有效鑒別。而對于主要靠雜交育種及種子園等有性繁殖的落葉松來說,優良家系及雜種的鑒別十分重要。本研究的試驗材料是從針葉中提取DNA,方法簡單、容易操作,且適用于落葉松雜種的早期選擇。在試驗過程中,對記錄數據結果的引物重復試驗了12次,結果穩定可靠,并在試驗后期對材料隨機編號,然后用特異性引物對其進行擴增,得到的結果與實際結果相符合。
本研究找到的合適的引物能夠對落葉松家系進行鑒別,可以為落葉松雜種優勢早期預測、雜種優勢機理研究提供技術支持。所有家系在0.4的遺傳距離上可以分成4種雜交組合,說明不同雜交組合的落葉松可以通過遺傳距離來進行區分。本研究表明,分子標記的結果可以真實、穩定地反映出家系間由于遺傳關系造成的差別。
[1]張含國,張成林,蘭士波,等.落葉松雜種優勢分析及家系選擇[J].南京林業大學學報:自然科學版,2005,29(3):69-72.
[2]羅旭,王祥岐,張含國,等.雜種落葉松的生長表現及遺傳增益[J].東北林業大學學報,2005,33(6):8-9.
[3]關麗鵬,王志剛,趙清風.雜種落葉松的識別技術[J].防護林科技,2006(5):89-90.
[4]趙墾田,張敩方,羅麗芬,等.長白落葉松種子活力與染色體核型[J].東北林業大學學報,1992,20(5):7-11.
[5]楊茜,喬辰,徐平,等.落葉松種子同工酶的研究[J].科學技術與工程,2005,5(12):797-799.
[6]Zietkiewicz E,Rafalski A,Labuda D.Genome fingerprinting by simple sequence repeat(SSR)-anchored polymerase chain reaction amplification[J].Genomics,1994,20:176-183.
[7]Liu Guifeng,Dong Jingxiang,Jiang Ying,et al.Analysis of genetic relationship in 12 species of Section Strobus with ISSR markers[J].Journal of Forestry Research,2005,16(3):213-215.
[8]Van Ripley L,Roslinsky V.Identification of an ISSR marker for 2-progenyl glucosinolate content in Brassica juncea L.and conversion to a SCAR marker[J].Molecular Breeding,2005,16(1):57-66.
[9]Arcade A,Anselin F,Rampant F P,et al.Application of AFLP,RAPD and ISSR markers to genetic mapping of European and Japanese larch[J].Theor Appl Genet,2000,100(2):299-307.
[10]Mehes M S,Nkongolo K K,Michael P.Genetic analysis of Pinus strobus and Pinus monticola populations from Canada using ISSR and RAPD markers:development of genome-specific SCAR markers[J].Pl Syst Evol,2007,267(1/4):47-63.
[11]張磊,張含國,李雪峰,等.落葉松種間及無性系間ISSR鑒別技術的研究[J].植物研究,2008,28(2):216-221.
[12]林萍,張含國,謝運海.正交設計優化落葉松ISSR-PCR反應體系[J].生物技術,2005,15(5):34-37.
[13]Sokal R R,Michener C D.A statistical method for evaluating systematic relationships[J].Univ of Kansas Sci Bulletin,1958,28:1409-1438.
[14]管玉霞.落葉松部分cpDNA,mtDNA及ITS序列研究及其應用于落葉松種的鑒定的可行性[D].北京:北京林業大學,2006.
[15]曲麗娜.興安、長白及華北落葉松RAPD和ISSR分子標記的物種特異性鑒定[D].哈爾濱:東北林業大學,2006.
[16]楊玉玲.杉木種質資源遺傳分析及無性系、雜交種ISSR指紋鑒定[D].福建:福建農林大學,2006.
