晉北鹽堿化賴草草地群落特征對不同放牧強度的響應
2011-03-14 09:10:14謝開云董寬虎王妍君
草業科學 2011年9期
謝開云,趙 祥,董寬虎,王妍君
(山西農業大學動物科技學院草地研究所,山西 太谷 030801)
草地牧草資源是放牧家畜的主要食物來源,草地放牧系統可持續發展的前提條件就是草地的健康或不退化[1],而放牧家畜的種類、合理的載畜量、適宜的放牧強度、正確的放牧時期和適當的放牧或刈割頻率是維系草地群落穩定性以及高的草地生產力的根本保障[2]。長期以來,草原草畜平衡問題倍受關注,不同類型草地合理載畜量一直是研究和生產人員關注的重點。關于不同載畜量對草地群落特征的影響,在草甸草原、典型草原、荒漠草原及高寒草甸草地進行了大量研究[3-6],主要是針對在不同放牧梯度上植物多樣性變化過程的研究[7]。研究表明,隨著載畜量的增加,一些適口性好、再生性強、耐牧的牧草種類減少,而適口性差、不耐牧的種類增多[8],導致草地植物生產力下降、多樣性降低、群落發生退化演替[9]。普遍支持中等載畜量條件下草地植物群落具有最高的植物多樣性,重度放牧造成群落多樣性減少、物種豐富度降低的觀點[10-11]。對鹽堿化草地載畜量的研究報道較少[12],另外中等載畜量條件是相對草地類型而言的,不同類型的草地由于草層高度和草地質量不同,其最適載畜量也不同。因此本研究針對鹽堿化賴草草地設置不同放牧強度,通過調查草地植物群落特征的變化來確定鹽堿化草地上合理的載畜量,為進一步優化草地的利用方式和利用率奠定基礎。
1 材料與方法
1.1自然概況 試驗地位于右玉縣威遠鎮后所堡村,40°10′48″ N,112°19′48″ E,海拔1 348 m,年均氣溫4 ℃,最冷月1月氣溫-15~-11 ℃,最熱月7月氣溫19~20 ℃,≥0 ℃積溫2 600~3 600 ℃·d。終霜期在5月初,初霜期為9月上、中旬,無霜期100~120 d。全年太陽總輻射量598 kJ/cm2,年日照時數2 600~2 700 h,7、8月份光能資源最豐富;年降水量450 mm,屬溫帶氣候。草地類型屬溫性草甸草原類山地草甸草原亞類,草地型為賴草(Leymussecalinus)-鵝絨委陵菜(Potentillaanserina)-雜類草群叢,主要生長有賴草、鵝絨委陵菜、艾蒿(Artemisialavandulaefolia)、米蒿(A.dalailamae)、豬毛菜(Salsolacollina)、草地風毛菊(Saussureaamara)、狗尾草(Setariaviridis)、堿茅(Puccinelliafloorida)、草木樨(Astragalusmelilotoides)、西伯利亞蓼(Polygonumsibiricum)、蘆葦(Phragmitescommunis)等。土壤為淡栗鈣土,土壤pH值為9.2。圖1a、b分別為試驗期間樣地降水量和氣溫的變化。
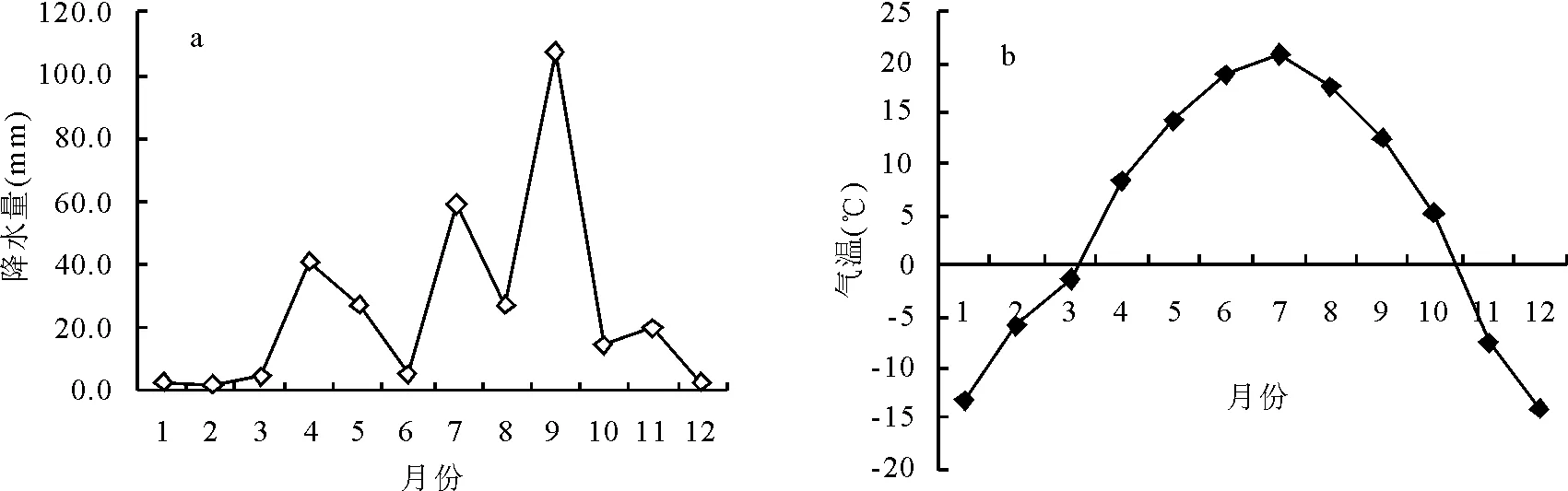
圖1 試驗期間樣地降水量和氣溫的變化
1.2供試動物 放牧動物為體質量(25±2.5)kg、健康的1歲當地綿羯羊20只,試驗前進行編號、藥浴、驅蟲。分組進行草地圍欄放牧。
1.3放牧試驗設計 放牧地的總面積2.25 hm2,并用圍欄分隔成15個小區,小區面積0.15 hm2,設不放牧(CK)、輕度放牧(LG)、中度放牧(MG)、重度放牧(HG)、極度放牧(EG)5個處理,放牧前測定草地鮮草產量,根據草地牧草利用率CK為0、LG為20%~35%、MG為40%~50%、HG為55%~65%、EG為70%~80%確定,其相應實際載畜量分別為4.44(LG)、8.88(MG)、13.32(HG)和17.76(EG)羊單位/(hm2·d),3個重復,每5個小區劃為一組,分別按CK、LG、MG、HG、EG 20只綿羊不放牧、放牧1 d、放牧2 d、放牧3 d、放牧4 d,10 d后放牧第2組小區,再過10 d放牧第3組小區,30 d為一個放牧周期。放牧時間為6月27日-9月2日,期間除下雨延誤不放牧時間外,2個放牧周期共約60 d。
1.4測定指標與方法 在輪牧各小區隨機布置5個活動圍籠(0.6 m×0.6 m),分別測定牧前(籠內)和牧后(籠外)牧草地上現存量。在各小區分別以“S”形布置10個固定樣方(1 m×1 m),于每輪放牧結束測定植物群落蓋度、頻度、密度、高度。蓋度采用點測法測定;頻度以1 m×1 m樣方測定,記錄某植物種的出現頻率;密度采用記錄單位面積的株數法測定;高度則隨機選取10株用卷尺測定,取平均值。
1.5數據處理 草地群落優勢度計算公式:
IV=(RH+RC+RF+RD)/4。
式中,IV為優勢度;RH為相對高度;RC為相對蓋度;RF為相對頻度;RD為相對密度。
群落α多樣性指數采用以下公式計算:
Margalef 物種豐富度指數:
Dma=(S-1)/lnN;
Shannon-Wiener 多樣性指數:
H=-ΣPiln(Pi);
Simpson 優勢度指數:
C=1-Σ(Pi)2;
E. Pielou 均勻度指數:
EPi=H/lnS。
式中,S為群落物種數目;N為所有物種個體總數;Pi為相對重要值。
所有數據采用SAS 9.0軟件進行差異顯著性分析和多重比較,Microsoft Excel繪圖。
2 結果與分析
2.1植物群落特征對不同載畜量的響應 經調查草地植物群落由13種主要植物組成,其中禾本科有賴草、蘆葦、堿茅和狗尾草4種,豆科只有草木樨1種,菊科有蒲公英(Taraxacummongolium)、米蒿、草地風毛菊和艾蒿4種,其余有薔薇科的鵝絨委陵菜、蓼科的西伯利亞蓼、藜科的豬毛菜和車前科的車前(Plantagoasiatica)等植物。
兩輪放牧以后,草地群落主要物種的蓋度、頻度、密度和高度均發生明顯的變化(圖2)。第1輪放牧后,隨放牧強度的增加,賴草、堿茅和西伯利亞蓼的蓋度減小(圖2a),其中賴草的蓋度不放牧時最大,為89.00%,在EG時最小,為60.67%,與不放牧相比減小31.83%;狗尾草和蒲公英先增加后減小;鵝絨委陵菜蓋度在CK時明顯高于LG、MG、HG、EG,但在LG、MG、HG和EG之間變化趨勢不明顯。賴草的頻度變化趨勢不明顯,鵝絨委陵菜、蒲公英和西伯利亞蓼的頻度先增加后減小,MG時最大,其他植物種頻度都減小(圖2c)。賴草的密度明顯高于其他物種(圖2e),隨放牧強度的增加先增加后減小,其中在CK和LG時值最大,HG時最小,其他植物種的密度變化趨勢不明顯。賴草高度除EG外,隨放牧強度增加而降低(圖2g),CK時值最大,為29.61 cm,HG時值最小,為21.34 cm,與CK相比減幅為27.92%;堿茅高度先增加后減小,LG時最大,其他植物高度變化規律不明顯。
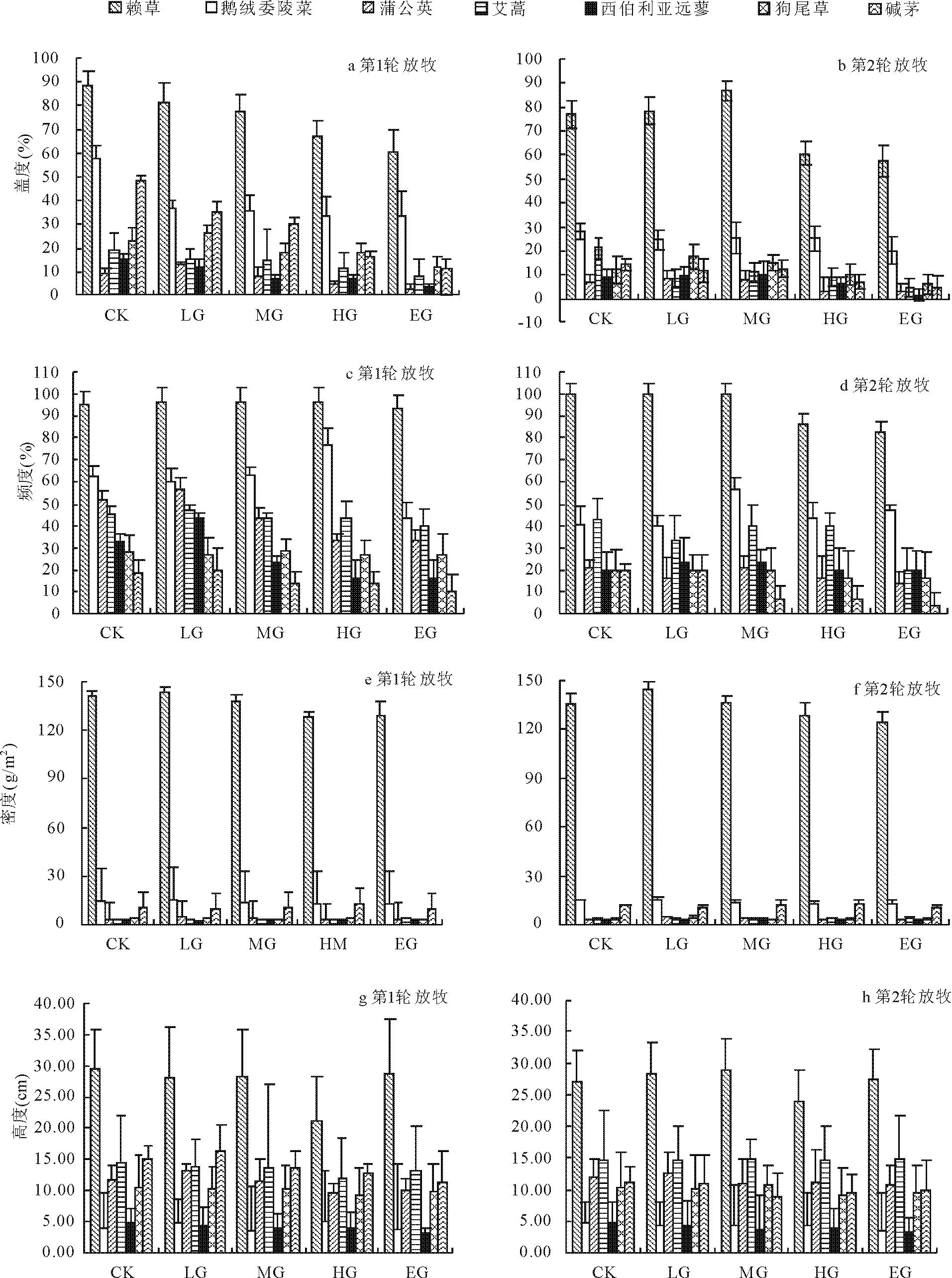
圖2 不同放牧強度對群落物種蓋度、頻度、密度和高度的影響
在第2輪放牧以后,隨放牧強度增加,賴草、狗尾草、蒲公英和西伯利亞蓼蓋度先增大后減小(圖2b),其中賴草MG時值最大,為86.67%,EG時值最小,為57.67%;堿茅和艾蒿蓋度減小;鵝絨委陵菜蓋度不放牧時最高,EG時最小。賴草的頻度在CK、LG、MG時變化不明顯,但在HG、EG時明顯下降;狗尾草、堿茅和西伯利亞蓼頻度先增加后減小;蒲公英的頻度值在CK和MG時最大;艾蒿在CK、MG,HG時較大,在EG最小。優勢物種賴草的密度LG時最大,EG時最小,其他植物種的密度變化趨勢不明顯。賴草的高度先增加后減小再增加,MG時最大,為28.84 cm,HG時最小,為23.98 cm,與MG相比減幅為16.85%;蒲公英在LG時最大,其他植物的高度變化趨勢不明顯。
兩次輪牧草地現存量的變化趨勢相同,第2輪放牧后的現存量略高于第1輪放牧后(圖3)。第1輪放牧后,在CK時最大,為2 473.4 kg/hm2,MG時較大,為2 354.2 kg/hm2,EG時最小,為1 496.0 kg/hm2,與CK、MG相比減幅分別為39.51%和36.45%。第2輪放牧后,在CK時最大,為2 678.4 kg/hm2,MG時較大,為2 594.7 kg/hm2,EG時最小,為1 802.6 kg/hm2,與CK、MG相比減幅分別為32.69%和30.52%。綜合所述,草地現存量隨著放牧強度的增大逐漸減小,但在中度放牧干擾下有增大的趨勢。
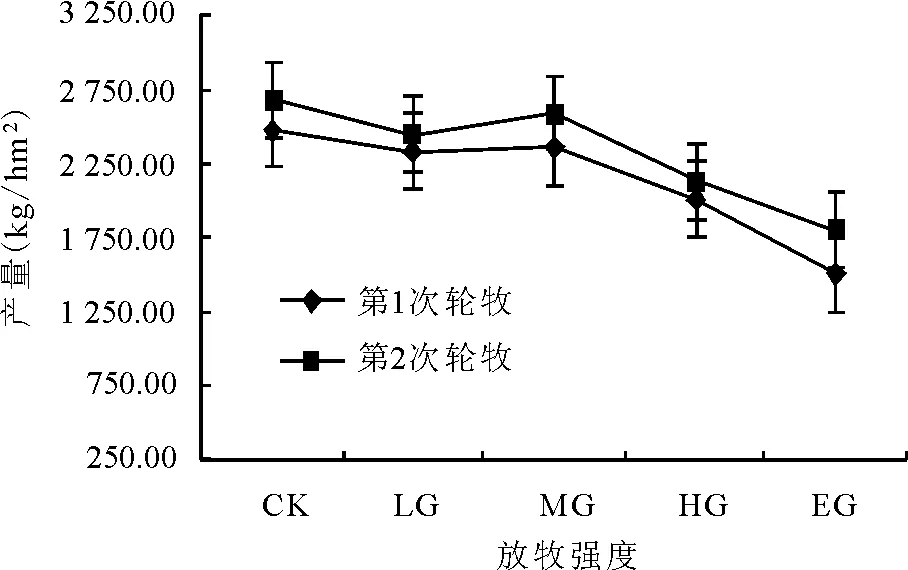
圖3 不同載畜量對草地現存量的影響
2.2植物優勢度對不同放牧強度的響應 隨著放牧強度的變化,群落的變化主要反映在不同植物種在群落中作用大小的消長上。隨著放牧強度的增大,賴草和鵝絨委陵菜優勢度都呈降低趨勢(表1)。第1輪放牧后,與不放牧的草地相比,賴草優勢度在LG增加了0.03%,MG、HG和EG分別降低了0.90%、1.20%、3.87%,鵝絨委陵菜的優勢度分別降低了2.83%、2.24%、2.47%、3.42%。第2次輪牧后,與不放牧草地相比,賴草的優勢度在LG和MG分別增加了1.21%和3.88%,HG和EG分別減低了0.66%和0.94%;鵝絨委陵菜的優勢度在LG和EG分別降低了1.27%和0.25%,MG和HG時分別增加了3.04%和2.15%。兩次輪牧后,隨著放牧強度的增大,米蒿、蒲公英、豬毛菜和西伯利亞蓼的優勢度都呈下降趨勢;堿茅的優勢度先增加后減小。另外MG時,草地風毛菊、鵝絨委陵菜和堿茅的優勢度較高,說明中度放牧強度有利于這3種草的生長。
2.3不同放牧強度下植物群落α多樣性的變化 不同放牧強度下群落物種多樣性指數分析(表2)表明,植物群落的Margalef物種豐富度指數MG顯著顯著高于CK、LG、HG和EG(P<0.05),Shannon-Wiener多樣性指數CK顯著高于LG、MG、HG和EG(P<0.05),Simpson優勢度指數MG和CK、LG之間差異不顯著(P>0.05),但顯著高于HG和EG(P<0.05),E.pielou均勻性系數MG和LG間差異不顯著(P>0.05),但二者都顯著低于HG和EG(P<0.05),這表明不放牧草地保持較高的物種多樣性,隨著放牧強度的增加,家畜對牧草采食強度增加,物種多樣性也隨之降低,中度放牧強度時群落物種具有較高的物種豐富度和優勢度,過度放牧下,家畜的采食量增加,降低了首先被選擇并連續采食的優勢物種,使優勢物種的優勢度減少。綜合所述,在適度放牧條件下,草地群落具有較高的物種豐富度和優勢度,但均勻度較低。
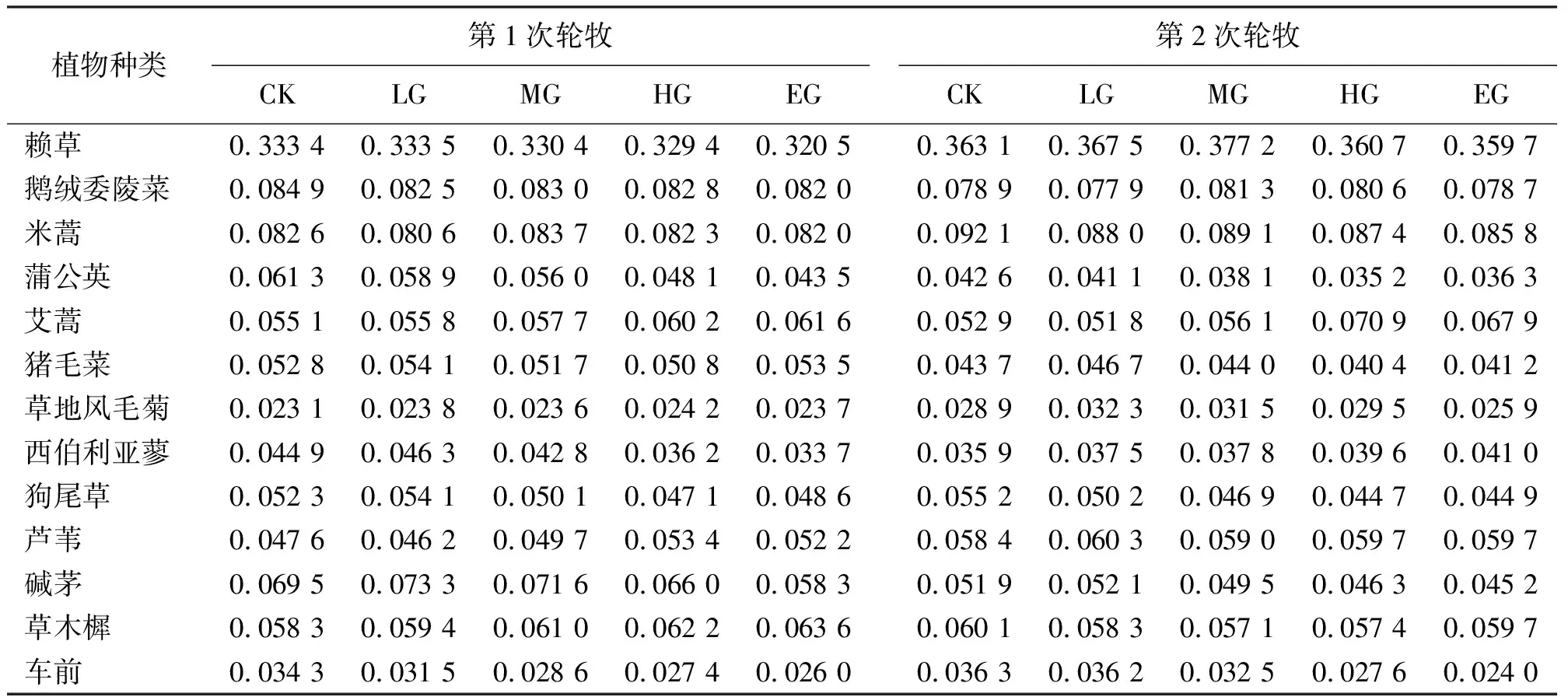
表1 不同放牧強度下賴草草地主要植物優勢度的變化
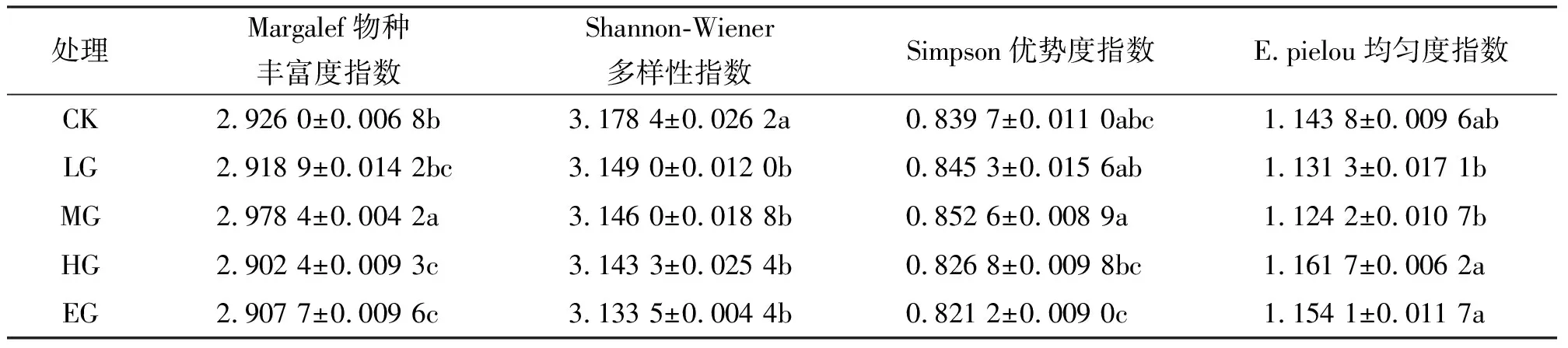
表2 不同放牧強度對豐富度指數、群落多樣性、優勢度指數和均勻度指數的影響
3 討論與結論
3.1不同放牧強度對草地群落特征的影響 放牧對草地植被的數量特征有著重要的影響,并且這種影響程度是隨著放牧強度增加而增強的,表現為隨著放牧強度的增加,草地植被地上生物量下降,蓋度、高度降低。通過兩次輪牧后,草地群落蓋度和頻度在中度放牧時較大,極度放牧時較小。比較兩次輪牧后的草地群落物種蓋度發現,主要優勢植物賴草在第1次輪牧后隨著放牧強度的增加而減小,而第2次輪牧后的變化趨勢為先增加后減小,這是由于兩次輪牧間隔時間只有30 d,第1輪放牧后賴草的蓋度變化主要是隨著放牧強度增加,放牧家畜采食總量增加,進而影響了賴草蓋度變化;而第2次輪牧后賴草蓋度變化是因為受到兩次放牧疊加效應的影響,表現出補償性生長的結果。兩次放牧后堿茅蓋度變化與其他植物種不同,隨著放牧強度的增加而減小,主要是由于7月和8月堿茅處在果后營養期,此季節堿茅生長緩慢,對放牧比較敏感,堿茅的最佳放牧利用時間應在春季(4月下旬-5月上旬),過早和過遲放牧都不利于堿茅的生長[15]。
在草地的實際利用與管理過程中,最簡單而常用的參數是植被群落或植物的高度與密度,特別是高度可以作為草地群落的表面特征參數(surface characteristic)[16]。放牧家畜作用于草地,最直接最明顯的變化就是草地物種高度的變化,家畜放牧能降低草地植物的高度,特別是優勢植物的高度[17]。草地是由多種植物構成的群落,由于放牧家畜對植物的喜好程度或偏食性的差異,植物高度的變化程度并不一致,從而導致草地植物高度變化的差異[17]。隨著放牧強度的變化,植物密度的變化不明顯,其原因可能是植物密度變化需要一個長期的過程,短時間放牧對植物密度影響不大。
隨著牧草生育期的推進,牧草的產量增加但品質卻下降;草地現存量高峰出現在極度放牧和不放牧,說明隨著放牧強度的增加,家畜采食總量增大,造成現存量的下降,但適度的放牧又會刺激植物的生長[18],因此極度放物時草地現存量高于其他放牧處理,這與韓文軍等[19]研究過度放牧對羊草雜草類群落種的構成和現存生物量的影響得出中度放牧區現存量高于重度放牧區而低于輕度放牧區的結論不同。這主要是由于草地在放牧干擾下所受放牧時間的不同而表現出不同的特征。
綜合所述,在經過兩輪放牧以后,鹽堿化賴草草地群落特征發生了明顯的變化,但兩次輪牧后草地的群落特征變化規律不同,其主要原因是兩輪放牧分別在7、8月進行,說明草地群落特征不僅受到家畜采食的影響也要受生長季節的影響。在同一輪放牧下,不同植物數量特征也呈現不同變化規律,這一方面與植物對放牧的耐受程度有關,另一方面與家畜對植物的喜好程度或偏食性有關。
3.2不同放牧強度對草地植物優勢度的影響 植物群落優勢度是評價植物種群在群落中作用的一項綜合性數量指標,在草地植被特征值的評價中得到廣泛應用,是應用最廣的物種特征值[20]。草地群落植物優勢度變化是種群間及其與環境之間的相互作用的反映,在一定程度上表征了在不同放牧強度下植物群落特征的變化,賴草的優勢度隨放牧強度的增加呈降低趨勢,這說明在草地群落的眾多影響因子中,放牧強度是重要的影響因子。
對試驗草地調查發現,該草地物種組成比較簡單,其優勢物種主要有賴草和鵝絨委陵菜,隨著放牧強度的增大,賴草和鵝絨委陵菜優勢度呈下降趨勢,說明賴草在首先被選擇并連續采食的情況下很難恢復,生長速度明顯降低,但下降的幅度比較小,這可能是試驗年限較短的緣故。在適度放牧下,禾本科類牧草在此類草地上屬相對比較耐牧的物種,在群落中其優勢度有增加的趨勢。這與張繪芳等[13]的研究,適度的放牧有利于草地植被結構的改善和優良牧草比例的提高,而過度放牧使草地禾本科牧草的比例降低,雜草的比例升高結果一致。另外,在群落中唯一一種匍匐型植物種鵝絨委陵菜的特征隨著放牧強度的增加也表現出減小的趨勢,一方面這可能與本試驗年限短有關,另一方面是由家畜踐踏而導致的。而彭祺和王寧[14]研究表明,高強度放牧下,草地植物生態型由直立型向匍匐型發展,這種放牧強度對草地植被結構的影響是隨著放牧強度的增加而加強的。
綜合所述,在中度放牧時,主要優勢種賴草的蓋度、頻度、高度和現存量都達到最高,保持其在草地中的優勢地位,說明中度放牧強度有利于草地優勢植物的生長和繁殖,同時可以初步確定該類鹽堿化草地的適宜載畜量為8.8羊單位/(hm2·d),但還需要進一步研究確定優化最適載畜量。
3.3不同放牧強度對草地群落α多樣性的影響 α多樣性是對一個群落內植物分布的數量和均勻程度的測量指標,是生物群落在組成、結構、功能和動態方面表現出的差異,反映了植物對環境的適應能力和對資源的利用能力。群落植物的豐富度指數反映出群落內植物種類的多少;多樣性指數顯示群落種群變化過程與環境的關系,Shannon-Wiener多樣性指數可表征由生物群落等級特征引起的多樣性程度;群落均勻度是指群落中各種類個體數量的分配比例,群落均勻度和群落優勢度是2個相反的概念。在本研究中,極度放牧下Margalef物種豐富度指數最大,表明適度的干擾可以增加群落的植物種類,進而影響草地的生產力和穩定性;不放牧的Shannon-Wiener多樣性指數顯著大于輕度放牧、極度放牧、重度放牧和極度放牧下(P<0.05),表明過度放牧干擾則會導致群落多樣性降低,與適度放牧可以降低群落優勢種排斥其他物種的能力,從而提高了群落水平上物種的多樣性[21]的結果不同,但與蒙旭輝等[22]對不同放牧強度下羊草草甸草原群落特征及多樣性結果一致,這可能與草甸草原類草地生長條件較好有關。中度放牧下群落優勢度指數顯著大于輕度放牧和極度放牧(P<0.05),而均勻度指數顯著低于輕度放牧和極度放牧(P<0.05),這與金曉明和韓國棟[23]分析放牧對草甸草原植物群落結構及多樣性的影響的結果不同,這可能是放牧時間的長短差異造成的,說明在適度放牧下群落優勢種明顯,優勢種的個體數和蓋度明顯高于一般種而群落均勻度降低[24]。群落物種的豐富度和多樣性程度越大,物種對生境的分割程度越高,因而群落物種均勻度降低[25]。群落在中度放牧強度下,物種保持較高的豐富度,但均勻度指數比較低。適度的放牧刺激了植物本身具有的補償性生長機制,放牧干擾對草地植物群落物種多樣性的影響符合“中度干擾理論”。
[1]宮海靜,王德利.草地放牧系統優化模型的研究進展[J].草業學報,2006,15(6):1-8.
[2]董全民,趙新全,馬玉壽,等.不同牦牛放牧率下江河源區垂穗披堿草/星星草混播草地第一性生產力及其動態變化[J].中國草地學報,2006,28(3):5-15.
[3]李永宏,汪詩平.放牧對草原植物的影響[J].中國草地,1999,7(3):11-19.
[4]趙寶山,王健.草場不同程度的利用對植被的影響[J].內蒙古草業,2000(4):21-24.
[5]楊剛,王思成,楊智明.不同放牧強度對荒漠草原植被和灘羊生產性能影響研究[J].中國飼料,2008(22):12-15.
[6]董全民.放牧強度對高寒混播草地群落結構的影響[J].青海畜牧獸醫雜志,2007,37(2):1-3.
[7]姚愛興,王培,樊奮成,等.不同放牧處理下多年生黑麥草/白三葉草地第一性生產力研究[J].中國草地,1998,24(2):12-16.
[8]董全民,趙新全,馬玉壽.放牧強度對混播草地群落數量特征的影響[J].草地學報,2007,15(4):394-397.
[9]陳佐忠,汪詩平.中國典型草原生態系統[M].北京:科學出版社,2000:9-15.
[10]Grimes J P.Control of species diversity in herbaceous vegetation[J].Journal of Environmental Management,1973,1:151-167.
[11]Huston M.A general hypothesis of species diversity[J].American Naturalist,1979,113:81-101.
[12]尚宗波.松嫩平原鹽堿化草地模擬模型研究[D].北京:中國科學院研究生院,2001.
[13]張繪芳,李霞,王建剛,等.塔里木河下游植物群落結構特征分析[J].生態環境,2007,16(4):1219-1224.
[14]彭祺,王寧.放牧與草地植物之間的相互關系[J].寧夏農學院學報,2004,25(4):76-79.
[15]趙明清,任憲濤,王東梅,等.朝鮮堿茅放牧留茬高度實驗[J].吉林農業科學,2009,34(5):45-49.
[16]Hodgson J.Variations in the surface characteristics of the sward and the short term rate of herbage in take by calves and lambs[J].Grass and Forage Science,1981,36(1):49-57.
[17]王德利,滕星,王涌鑫,等.放牧條件下人工草地植物高度的異質性變化[J].東北師大學報(自然科學版),2003,35(1):102-109.
[18]張謐,王慧娟,于長青.珍珠草原對不同模擬放牧強度的響應[J].草業科學,2010,27(8):125-128.
[19]韓文軍,春亮,侯向陽.過度放牧對羊草雜草類草群落的構成和現存生物量的影響[J].草業科學,2009,26(9):195-199.
[20]汪詩平,李永宏,王艷芬,等.不同放牧率對內蒙古冷蒿草原植物多樣性的影響[J].植物學報,2001,43(1):89-96.
[21]沈彥,張克斌,杜林峰,等.封育措施在寧夏鹽池草地植被恢復中的作用[J].中國水土保持科學,2007,5(3):90-93.
[22]蒙旭輝,李向林,辛曉平,等.不同放牧強度下羊草草甸草原群落特征及多樣性分析[J].草地學報,2009,17(2):240-244.
[23]金曉明,韓國棟.放牧對草甸草原植物群落結構及多樣性的影響[J].草業科學,2010,27(4):7-10.
[24]曹成有,蔣德明,阿拉木薩,等.小葉錦雞兒人工固沙植被恢復生態過程的研究[J].應用生態學報,2000,11(3):349-354.
[25]Reynolds C M,Wolf D C.Effect of soil moisture and air relative humidity on ammonia volatilization from surface-applied urea[J].Soil Science,1987,143:144-152.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
趣味(語文)(2020年3期)2020-07-27 01:42:46
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
