大菱鲆早期發育過程中免疫器官的發生
2011-01-11 14:41:32佟雪紅徐世宏劉清華肖志忠馬道遠
海洋科學 2011年6期
佟雪紅, 徐世宏, 劉清華, 李 軍, 肖志忠, 馬道遠
(1. 中國科學院 海洋研究所, 山東 青島 266071; 2. 鹽城師范學院, 江蘇 鹽城 224002; 3. 中國科學院 研究生院, 北京 100039)
大菱鲆早期發育過程中免疫器官的發生
佟雪紅1,2,3, 徐世宏1, 劉清華1, 李 軍1, 肖志忠1, 馬道遠1
(1. 中國科學院 海洋研究所, 山東 青島 266071; 2. 鹽城師范學院, 江蘇 鹽城 224002; 3. 中國科學院 研究生院, 北京 100039)
應用組織切片研究了大菱鲆(Scophthalmus maximus)初孵仔魚至 60日齡幼魚免疫器官的發育過程。結果表明, 免疫器官原基出現的先后順序是頭腎、脾臟和胸腺。1日齡仔魚可以觀察到頭腎原基, 包含未分化的造血干細胞。5日齡時, 脾臟原基出現, 其淋巴化開始于27日齡且發育速度較慢。13日齡時, 大菱鲆仔魚胸腺原基出現, 且發育速度較快, 分為外區和內區。在大菱鲆早期發育過程中, 胸腺和頭腎之間出現細胞遷移現象。免疫器官淋巴化的順序是胸腺、頭腎和脾臟。在免疫器官發育后期, 脾臟和頭腎中均發現了黑色素-巨噬細胞中心(MMCs), 脾臟中較豐富, 但在胸腺中尚未發現。在大菱鲆仔魚早期發育階段, 免疫器官尚未發育完善, 非特異性防御機制起著重要的免疫作用。
大菱鲆(Scophthalmus maximus); 早期發育階段; 免疫器官; 發生; 組織學
魚類免疫系統是機體識別和消除“異物”的防衛系統, 是魚類防止病原入侵的第一防線[1]2,[2]649。參與魚類免疫應答的免疫器官主要是腎臟、脾臟、胸腺, 它們是魚類免疫細胞發生、分化、成熟和增殖的主要場所。因此, 研究魚體免疫器官的個體發生, 是深入認識魚體免疫機制建立的基礎, 對提高魚苗的培育技術有重要的應用價值。目前, 國內對鲇魚(Silurus asotus)[3], 斜帶石斑魚(Epinephelus coioides)[4]和大黃魚(Pseudosciaena crocea)[5]等免疫器官的發生發育規律進行了探討。而對于鲆鰈魚類免疫系統的研究, 主要見于Chantanachookhin等[6]146和 Liu等[7]對牙鲆(Paralichthys olivaceus)的研究、Padrós 等[1]2對大菱鲆(Scophthalmus maximus)的研究、Bowden等[8]對大西洋庸鰈(Hippoglossus hippoglossus)的報道及肖志忠等[9]88對條斑星鰈(Veras per moseri)的研究。
大菱鲆屬鲆科(Bothidae)、菱鲆屬(Scophthalmus),自然分布于東北大西洋沿岸, 其生長迅速、適應低水溫生活, 市場潛力巨大。中國自1992年首次引入以來, 經過十幾年的發展, 大菱鲆的工廠化養殖已成為中國一項新興養殖產業, 有效推動了中國海水養殖第四次浪潮的形成和發展[10-12]。然而, 大菱鲆養殖產業也面臨諸多挑戰, 魚苗的高畸形率和低成活率仍然是生產實踐中的難點問題, 推測部分原因可能與其免疫組織尚未完善以及其免疫功能相對滯后有關。要解決這一問題, 需要研究魚類免疫系統的早期發生, 搞清楚魚類胚胎、仔稚魚和幼魚免疫能力及其免疫系統成熟機制[13]。作者從免疫器官發育方面尋找產生這些問題的可能性因素, 旨在為促進大菱鲆養殖業的健康發展提供理論指導。1
1 材料與方法
實驗中大菱鲆魚苗取自山東煙臺東方海洋科技股份有限公司。仔魚的培育密度是125 尾/升。培育時的水質條件是: 水溫18-19 °C, 溶氧7.0-8.5 mg/L,鹽度30-32, pH 7.7-8.0。魚苗孵化出膜后至40日齡,每天取樣1次; 40日齡以后每2 d 取樣1次, 連續取樣60 d, 取樣在早晨投喂之前進行。樣品用Bouin’s液固定, 石蠟包埋, 用KD1508A輪轉式切片機(浙江金華儀器有限公司生產)連續切片, 分別進行橫切(從頭到尾)、縱切(從體左側到右側)和水平切(從背到腹)方向連續切片, 切片厚度為4-7 μm, H. E染色, 中性樹膠封片, NIKON SMZ-800顯微鏡下觀察, 照像。
2 結果
2.1 腎臟的發生
1日齡仔魚(2.82 mm), 腎為1對簡單的直腎管,位于卵黃囊和脊索之間, 頭腎間組織出現未分化的干細胞(圖 1-1)。2日齡仔魚(2.90 mm), 腎臟向前延伸, 腎管分支形成前腎管; 向后腎管沿著脊椎直抵軀干后部, 開口于肛門。3日齡仔魚(3.29 mm), 前腎管區間開始出現前腎小管, 但不發達。5日齡仔魚(3.62 mm), 前腎小管增多, 管壁內可見一些被伊紅染成粉紅色的分泌細胞, 在前腎管之間可見干細胞。7日齡時(3.91 mm), 前腎小管中出現分泌細胞, 腎間組織出現許多血細胞, 同時干細胞也增多(圖1-2)。8日齡時(4.1 mm)腎間組織出現淋巴母細胞。14日齡時(5.46 mm), 在頭腎部位的腎管區可見活躍的淋巴細胞生成(圖1-3)。
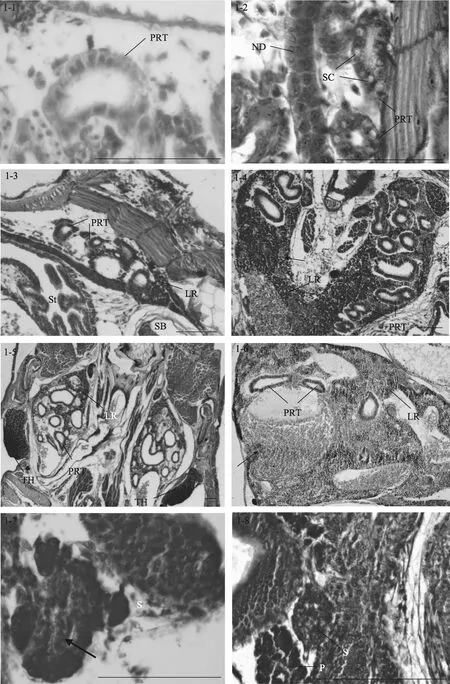
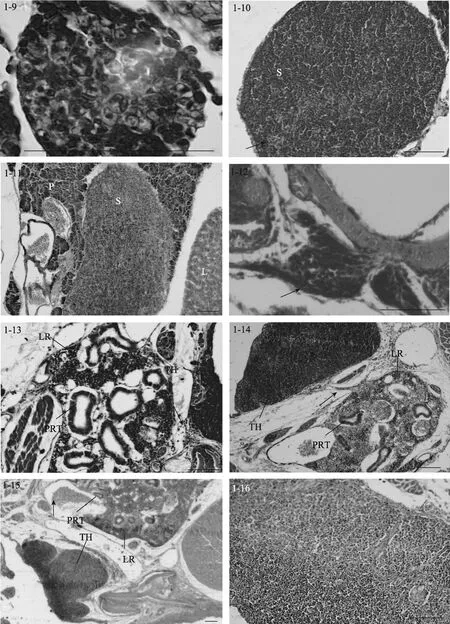
圖1 大菱鲆免疫器官的發育Fig. 1 Development of immune organs in turbot
19日齡時(7.17 mm), 淋巴細胞體積變小, 染色變深, 預示著頭腎淋巴化的開始。22日齡時(8.62 mm), 頭腎組織體積變大, 前腎小管數量較多, 管間組織包含大量已淋巴化的小淋巴細胞和紅細胞, 同時出現許多血竇(圖 1-4)。27日齡時(14.05 mm), 腎管區間有發達的血管, 血管內有豐富的紅細胞(圖1-5)。32日齡時(17.22 mm), 頭腎淋巴造血組織比率增加, 前腎小管數量減少, 腎小管上皮細胞開始退化, 有的細胞輪廓和界限不清晰, 細胞的嗜酸性大為增加, 為伊紅深染。此時體腎組織由大量的腎管、淋巴細胞和紅細胞占據, 頭腎在結構上與體腎不同。
隨著生長發育的進行, 37日齡時(20.59 mm)頭腎組織的血管周圍出現黑色素細胞, 并形成黑色素-巨噬細胞中心(melano-macrophage centers,簡稱MMCs)。50日齡時(24.76 mm), 頭腎組織可見大量的血竇和血細胞, 腎小管數量明顯減少(圖1-6); 體腎中紅細胞密集, 所占比例增大。60日齡時(31.71 mm),除了紅細胞生成集落和各種淋巴細胞外, 頭腎組織中仍然可見殘存的前腎小管, 但結構退化, 此后頭腎逐漸失去了分泌和排泄的功能, 而成為具有免疫機能和造血功能的淋巴樣組織。體腎逐步成熟, 作為排泄器官, 體腎組織中出現大量的腎小管, 同時還存在豐富的血細胞和淋巴細胞, 說明體腎在造血和免疫方面也有一定的作用。
2.2 脾臟的發生
5日齡時(3.62 mm), 脾臟原基出現, 位于前腸和鰾之間, 呈橢圓形, 被胰臟組織包圍(圖1-7), 由疏松的間充質細胞索和毛細血管組成, 內含少量微嗜堿性細胞或造血干細胞。隨著仔魚的生長發育, 脾臟逐漸向腹部移動, 在 9日齡時(4.47 mm), 脫離胰臟的包圍(圖 1-8)。15日齡時(5.8 mm), 脾臟體積變大, 此時可見活躍的紅細胞生成, 嗜堿性細胞和紅細胞數量增多。同時成纖維細胞、嗜堿性細胞和紅細胞形成了疏松的細胞索(圖1-9)。
27日齡時(14.05 mm), 脾臟體積顯著增大, 嗜堿性細胞明顯增多且進一步分化, 體積變小, 預示了脾臟的淋巴化。同時可見許多微血管, 內含豐富的紅細胞, 并形成原始的脾竇(圖 1-10)。34日齡時(17.97 mm), 脾臟結構緊密, 脾竇數量增加。網狀細胞通過胞質擴張形成網狀組織。微血管發達, 淋巴細胞和紅細胞更為豐富, 此時紅細胞數量明顯占優勢,為優勢細胞。40日齡時(21.4 mm), 紅細胞和類紅細胞進一步增多, 血管密集。50至60日齡(24.76-31.71 mm), 脾臟的結構與成體類似, 脾臟的網狀內皮系統十分發達, 表明其具有造血和免疫功能。在脾竇附近可見大量含黑色素的巨噬細胞并形成 MMCs (圖1-11)。隨著魚體的生長發育, 脾臟的組織學變化很少,主要是細胞組織的進一步生長和脾臟體積的增大。
2.3 胸腺的發生
13日齡時(5.4 mm), 大菱鲆仔魚中胸腺原基出現, 為兩側對稱的1對器官, 位于鰓腔上角背側, 靠近鰓蓋骨的背關節(圖1-12)。胸腺腹面為一層扁平上皮與鰓腔分隔。胸腺由未分化的嗜堿性淋巴母細胞組成, 染色較深。17日齡時(6.7 mm), 胸腺明顯, 開始出現小淋巴細胞, 此時尚無內區(髓質)和外區(皮質)之分。23日齡時(9.2 mm), 胸腺和頭腎位置較近,可見細胞遷移現象, 胸腺結構變得緊密(圖1-13)。
隨著生長發育的進行, 在27日齡時(14.05 mm),胸腺外被一層較薄的、含分泌樣細胞的上皮, 并初步分區為內區和外區, 但此時內區和外區之間沒有明顯的界限。在31日齡時(16.92 mm), 胸腺中出現血管, 內含血細胞。內區和外區主要由淋巴細胞和網狀上皮細胞構成。內區部分染色較淺, 網狀上皮細胞和類肌細胞相對較多, 淋巴細胞大而染色淺、數量少且排列松散; 外區部分染色深, 淋巴細胞小而染色深、數量多且排列密集, 無結締組織。35日齡時(18.73 mm), 胸腺中出現許多血管, 包含血細胞, 內區和外區分區逐漸明顯。同時可見胸腺和頭腎組織間明顯的細胞遷移現象, 即連接頭腎與胸腺的細胞“橋”(圖 1-14)。40 日齡時(21.4 mm), 外區和內區的淋巴細胞分布密集, 內區之下主要由一些稀疏的結締組織構成, 結締組織形成被膜將胸腺分割成許多不完全的胸腺小葉, 每個小葉可分為外區和內區, 小葉間的外區組織互相連接(圖 1-15)。50-60日齡(24.76 mm-31.71 mm), 幼魚的胸腺與成魚相似, 外區主要由淋巴細胞和上皮細胞構成, 外被一層黏膜分泌樣細胞上皮層; 內區部分淋巴細胞稀疏而網狀上皮細胞較多, 同時可見大量的血管和血細胞延伸在胸腺實質中(圖1-16)。
3 討論
本實驗中, 大菱鲆淋巴器官原基出現的順序是頭腎、脾臟和胸腺, 這跟Padrós等[1]12在大菱鲆中得出的試驗結果一致, 也跟已報道的大多數海水魚類如金鯛(Sparus aurata)[14]40、鱸魚(Dicentrarchus labrax)[15,16]和大西洋鱈魚(Gadus morhua)[17]512等類似; 但與羅非魚(Apistogramma alacrina)[18]和鯉魚(Cyprimus carpio)[19]等淡水魚不同, 其免疫器官原基的出現順序為胸腺、頭腎和脾臟。大菱鲆免疫器官淋巴化的順序是胸腺、頭腎和脾臟, 這跟大多數硬骨魚類免疫器官淋巴化的順序一致。但比較特殊的是大西洋鱈魚, 其免疫器官淋巴化的順序是頭腎、脾臟、胸腺; 而條斑星鰈, 其首先淋巴化的器官是頭腎,然后是胸腺, 最后是脾臟; 推測原因可能是大西洋鱈魚(Gadus morhuaL)和條斑星鰈對冷水溫養殖環境長期適應的結果, 即水溫影響了免疫器官的發生和分化[9]90, [17]515。
本實驗中組織學試驗數據顯示, 在 23日齡時,大菱鲆胸腺和頭腎之間可見細胞遷移現象, 即連接頭腎與胸腺的細胞“橋”, 在金鯛等硬骨魚中也發現了類似現象[14]45。另外, 本研究中大菱鲆頭腎組織在1日齡的仔魚中就存在未分化的造血干細胞, 而胸腺在13日齡時出現, 說明頭腎的干細胞并非來自于胸腺; 而且細胞橋的存在證實了大菱鲆在早期發育過程中, 頭腎的造血干細胞會遷移到胸腺中, 控制胸腺的分化, 這跟Padrós等[1]12在大菱鲆中得出的試驗結果一致, 同時也支持了“頭腎和胸腺是同一種干細胞分化出的不同器官”的觀點[20]140,[21]78。
黑色素-巨噬細胞中心是存在于真骨魚類免疫器官如腎臟、脾臟及軟骨魚類肝臟中的一種結構[22]。研究表明, MMCs有儲存鐵血黃素的作用并參與了紅細胞的凋亡過程[23,24]。本試驗中, 大菱鲆在脾臟和頭腎中均發現 MMCs, 以脾臟中較為豐富, 但在胸腺中尚未發現此類細胞。但牙鲆和真鯛(Pagrosomus major)與此不同, 兩者腎臟中 MMCs比脾臟中豐富[6]147。造成MMCs形態和數量差異的原因, 可能跟物種和魚類的養殖環境及生態習性有關。另外, 魚類中MMCs通常在淋巴器官發育成熟后的幼魚階段出現, 本實驗的結果與此相類似; 但鮭鱒魚通常在仔魚開口時出現MMCs[25]。
大菱鲆在早期生活階段要經歷變態期, 魚苗的形態結構發生顯著改變, 器官發育尚未完善, 此時魚苗很脆弱、死亡率較高。組織學顯示, 大菱鲆淋巴器官的成熟要在變態完成后, 而免疫功能的發育完善要在幼魚階段。因此, 作者認為大菱鲆仔稚魚階段出現的較低成活率可能與其免疫器官尚未發育完善以及免疫功能相對滯后有關。在早期生活階段, 魚苗特異性免疫受到限制, 因此, 非特異性防御機制起著重要的免疫作用, 如: 巨噬細胞、粒細胞、溶菌酶以及網狀內皮系統等[6]145。上述非特異性細胞不僅對抗原有吞噬作用, 還具有處理抗原和將抗原決定簇呈遞給淋巴細胞的潛在功能, 對于早期階段特異性免疫的缺乏起了一定的補償作用[26-28]。另外, 母源性免疫因子在魚類生活早期也起著同樣重要的免疫保護作用[4]824,[20]136。通過母體免疫方法增強魚體先天性的抵抗能力, 以及通過對魚苗進行有效的免疫接種提高魚體自身的免疫力才能從根本上減少發病率,實現健康養殖。
[1]Padrós F, Crespo S. Ontogeny of the lymphoid organs in the turbotScophthalmus maximus: a light and electron microscope study[J]. Aquaculture, 1996, 144 .
[2]張永安, 孫寶劍, 聶品. 魚類免疫組織和細胞的研究概況[J]. 水生生物學報, 2000, 24(6) .
[3]鐘明超, 黃浙. 鲇魚淋巴樣器官的發育[J]. 水產學報,1995, 3(3): 258-262.
[4]吳金英, 林浩然. 斜帶石斑魚淋巴器官個體發育的組織學[J]. 動物學報, 2003, 49(6).
[5]徐曉津, 翁朝紅, 王軍, 等. 大黃魚早期發育過程中免疫器官的發生[J]. 海洋學報, 2007, 29(3): 105-113.
[6]Chantanachookhin C, Seikai T, Tanaka M. Comparative study of the ontogeny of the lymphoid organs in three species of marine fish[J]. Aquaculture, 1991, 99 .
[7]Liu Y, Zhang S C, J iang G L, et al . The development of the lymphoid organs of flounder,Paralichthys olivaceus, from hatching to 13 months [J]. Fish Shellfish Immun, 2004, 16: 621-632.
[8]Bowden T J, Cook P, Rombout J H. Development and function of the thymus in teleosts [J]. Fish Shellfish Immun, 2005, 19: 413-427.
[9]肖志忠, 于道德, 孫真真, 等. 條斑星鰈免疫器官個體發生的組織學觀察[J]. 海洋科學, 2008, 32(7) .
[10]雷霽霖, 劉新富. 大菱鲆引進養殖的初步研究[J]. 現代漁業信息, 1995, 10(11): 1-3.
[11]雷霽霖, 劉新富, 馬愛軍. 大菱鲆的引進與馴養實驗[M]. 北京: 中國林業出版社, 1998,: 408-413.
[12]雷霽霖, 馬愛軍, 劉新富, 等. 大菱鲆(Scophthalmus maximusL. )胚胎及仔稚幼魚發育研究[J]. 海洋與湖沼, 2003, 34(1): 9-19.
[13]張士璀, 李欣, 汲廣東. 魚類免疫系統的早期發生[J].中國海洋大學學報, 2003, 37(4): 557-562.
[14]Josefsson S, Tatner M F. Histogenesis of the lymphoid organs in sea bream (Sparus aurata, L.)[J]. Fish Shellfish Immun, 1993, 3 .
[15]Quesada J, Villena M I, Navarro V. Ontogeny of the sea bass spleen (Dicentrarchus labrax): A light and electron microscopic study[J]. J Morphol, 1994, 221: 161-176.
[16]Abelli L, Picchietti S, Romano N. Immunocytochemical detection of thymocyte antigenic determinants in developing lymphoid organs of sea bassDicentrarchus labrax[J]. Fish Shellfish Immun, 1996, 6: 493-505.
[17]Schroder M B, Villena A J, Jorgensen T O. Ontogeny of lymphoid organs and immunoglobulin producing cells in Atlantic cod (Gadus morhuaL.)[J]. Dev Comp Immunol, 1998, 22 .
[18]Fishelson L. Cytological and morphological ontogenesis and involution of the thymus in cichlid fishes(Apistogramma alacrina) [J]. J Morphol, 1995, 223:175-190.
[19]Romano N, Taverne-Thiele A J, Fanelli M. Ontogeny of the thymus in a teleost fish,Cyprimus carpioL, Developing thymocytes in the epithelial microenvironment[J]. Dev Comp Immunol, 1999, 23: 123-127.
[20]Nakanishi T. Ontogeny of the immune system inSebasticus marmoratus: Histogenesis of the lymphoid organs and effect of theymectomy[J]. Environ Biol Fish, 1991,30 .
[21]Peterman A E, Petrie H L. Ontogeny of American paddlefish lymphoid tissues[J]. J Fish Biol, 2006, 69:72-81.
[22]Agius C, Roberts R J. Effect of starvation on the melano-macrophage centres of fish[J]. J Fish Biol,1981, 19: 161-169.
[23]Agius C. The role of melano-macrophage centres in iron storage in normal and diseased fish[J]. J Fish Dis,1979, 2: 337-343.
[24]Herraez M P. Structure and function of the melano-macrophage centres of the goldfishCarassius auratus[J]. Vet Immunol Immunop, 1986, 12:117-126.
[25]Agius C. Preliminary studies on the ontogeny of the melano-macrophages of teleost haemopoietic tissues and age-related changes[J]. Dev Comp Immunol, 1981,5: 597-606.
[26]Watts M, Kato K, Munday B I, et al. Ontogeny of immune system organs in northern bluefin tuna (Thunnus orientalis, Temminck and Schlegel 1844)[J]. Aquac Res,2003, 34: 13-21.
[27]蘇友祿, 馮娟, 郭志勛, 等. 軍曹魚淋巴器官發育的形態學研究[J]. 海洋水產研究, 2008, 29(4): 7-15.
[28]肖克宇. 水產動物免疫與應用[M]. 北京: 科學出版社, 2007: 155-156.
Ontogeny of immune organs during early developmental stages of turbot
TONG Xue-hong1,2,3, XU Shi-hong1, LIU Qing-hua1, LI Jun1, XIAO Zhi-zhong1, MA Dao-yuan1
(1. Institute of Oceanology, the Chinese Academy of Sciences, Qingdao 266071, China; 2. Yancheng Teachers University, Yancheng 224002, China; 3. Graduate University of Chinese Academy of Sciences, Beijing 100039, China)
Jun., 14, 2010
turbot (Scophthalmus maximus); early developmental stage; immune organs; ontogeny; histology
A histological study was made for the development of immune organs in turbotScophthalmus maximusfrom hatching to 60 days after hatching (DAH). The sequence of immune organ anlages appearance were head kidney, spleen and thymus. At 1 DAH, head kidney anlage and primordial stem cells were observed. Spleen anlage was present at 5 DAH, it became lymphoid at 27 DAH and developed slowly. Progenitor thymus appeared at 13 DAH and grew quickly. An outer zone and an inner zone in the thymus were observed. Cell migration occurred between thymus and head kidney. The first functional lymphoid organ was thymus followed by head kidney and spleen.During the posterior developmental period of the immune organs, the melano-macrophage centers (MMCs) were found in spleen and head kidney, but not in thymus. The abundance in spleen was higher. During early developmental stages of turbot larvae, immune organs did not grow perfectly and non-specific systems may play an important role in the immunocompetence mechanisms.
S961.6
A
1000-3096(2011)01-0062-06
2010-06-14;
2010-11-15
現代農業產業技術體系建設專項資金資助項目(nycytx-50); 國家863“十一五”計劃資助項目(2006AA10A404)
佟雪紅(1979-), 女, 山東濰坊人, 博士, 主要從事海洋生物學研究, 電話: 0532-82898716, E-mail: txh113005@163.com; 李軍,通信作者, 研究員, E-mail: junli@ms.qdio.ac.cn
譚雪靜)
