不同受害類型松林中馬尾松毛蟲自然種群生命表研究
2010-11-22 08:00:36劉興平溫小遂吳自榮
環境昆蟲學報 2010年2期
劉興平,溫小遂,李 冬,吳自榮
(1.江西農業大學園林與藝術學院,南昌 330045;2.江西省林業有害生物防治檢疫局,南昌 330077)
當前,研究生態系統中“植物 -植食性昆蟲 -天敵”三營養層次的相互關系已成為國內外昆蟲生態學的研究熱點之一。昆蟲的取食能引起被害植物產生某些變化,并形成一定的耐害和抗蟲機制,這種抗蟲機制是植物與昆蟲長期協同進化的結果,也是害蟲生態調控的重要基礎(Karban and Baldwin,1997;婁永根和程家安,1997;Agrawal,1998)。我國南方各省區的主要造林樹種之一 -馬尾松 Pinus massoniana Lamb周期性地遭受馬尾松毛蟲 Dendrolimns punctatus Walker的危害。該蟲已經成為我國南方森林生態系統中最具危害性的森林害蟲之一。為此,眾多學者對該蟲進行了長期的研究。當前,在研究馬尾松與馬尾松毛蟲之間的相互關系時已經證實馬尾松在受到馬尾松毛蟲取食后,松針中的水分和營養物質下降,而次生物質(如單寧、酚酸等)的含量上升,從而能誘導受害的馬尾松產生一定的抗馬尾松毛蟲的作用(劉興平等,2003;王國紅等,2008)。自從 Morris應用生命表技術研究云杉卷葉蛾 Choristoneura fumiferana自然種群動態以來(Morris and Miller,1954),利用生命表技術來研究昆蟲自然種群動態和發展趨勢也已經成為評價寄主抗性的一種非常重要而有效的方法。然而,在自然條件下利用種群生命表的方法進行松樹抗耐蟲性評價尚未見報道。本文通過野外實地調查,比較了自然條件下馬尾松毛蟲在受害松林與未受害松林中種群數量的消長動態,并應用生命表法探討了受害松樹對馬尾松毛蟲種群生物學參數的影響,以期為松樹的誘導抗性研究以及對馬尾松毛蟲的生態調控提供理論依據。
1 材料與方法
1.1 試驗地概況
試驗地設在江西省新建縣生米鎮和望城鄉。兩地各選一塊標準地(面積為 20×30 m2),兩塊標準地均為馬尾松純林,平均樹高 1.5~2 m,樹齡均在 9~10年,林下植被裸露。其中,望城鄉的馬尾松林地在近 5年均未發生過馬尾松毛蟲災害,針葉保存完好;生米鎮的馬尾松純林被當年第一代馬尾松毛蟲危害過且針葉保存率僅有 30%~50%。我們把望城鄉的馬尾松純林作為未受害松林,生米鎮的馬尾松純林作為受害松林。同時,從受害林地中采集一定數量的卵人工接入未受害松林中,在受害松林中的卵為第一代馬尾松毛蟲成蟲自然條件下所產的卵,兩地同步進行調查。
1.2 自然種群生命表的組建
1.2.1 卵期調查:按東、西、南、北各方位取樣,在針葉上編號標記一定數量的卵,隔日觀察卵的死亡和孵化情況,分別記錄正常的、發育不良的、被寄生的卵的數量。
1.2.2 幼蟲期調查:在樣枝上套上紗網罩,分 1~2齡期、3~4齡期、5~6齡期 3個蟲期,隔天調查樣樹中幼蟲的死亡數和失蹤數,同時在林間設置 3 m×3 m×2 m的大籠罩 5個,與觀察區幼蟲同步分階段接入各齡幼蟲,觀察記載幼蟲的自然損失數,所得數據用于換算天敵的捕食作用。
1.2.3 蛹期調查:待樣樹上的松毛蟲結繭后,將蟲繭采回移入室內,3 d后將繭剖開,鑒別♀、♂并統計數量。待蛹羽化后,記錄蛹的自然死亡數及天敵寄生的蛹數。
1.2.4 成蟲期調查:將各樣地中的蟲繭采回并用剪刀剖繭取蛹放入室內養蟲籠中,讓其自然羽化。逐日統計成蟲羽化數,并按♀:♂=1:1配比放入養蟲籠中,統計其自然產卵量。
1.3 生物學參數分析
死亡率(%)=消亡蟲數/起始蟲數 ×100
種群趨勢指數 I(Index of population development trend)=Ni+1/Ni(Ni+1:下代卵量 ;Ni:當代卵量)
死亡因素致死力 Ki=lgLxi-lgLxi+1;K=K1+K2+K3+… +Kn(Lx:每個階段的存活蟲數;Ki:階段K值;K:世代總 K值)
2 結果與分析
2.1 種群趨勢指數
種群趨勢指數是研究種群數量動態的一個主要指標。通過對林間樣樹中的馬尾松毛蟲各發育階段的存亡數量及其致死因子的系統調查和研究、以及卵期、幼蟲期天敵寄生或捕食情況觀察與籠罩接蟲輔助觀察試驗所獲得數據的處理,按 Price(1975)的方法組建了受害馬尾松林和未受害馬尾松林中第 2代馬尾松毛蟲的自然種群生命表(表1)。從表 1可以看出,在未受害松林中馬尾松毛蟲的世代存活率和種群趨勢指數為 15.33%和24.0489,明顯高于受害松林(0.61%和 0.4864)。說明在未受害松林中馬尾松毛蟲的自然種群數量明顯高于受害松林。同時,馬尾松毛蟲在未受害松林中的種群趨勢指數 >1,而在受害松林中的種群趨勢指數 <1,即馬尾松毛蟲在未受害松林中的自然種群繁殖一代后種群數量呈增長趨勢,而在受害松林中其種群呈下降趨勢。
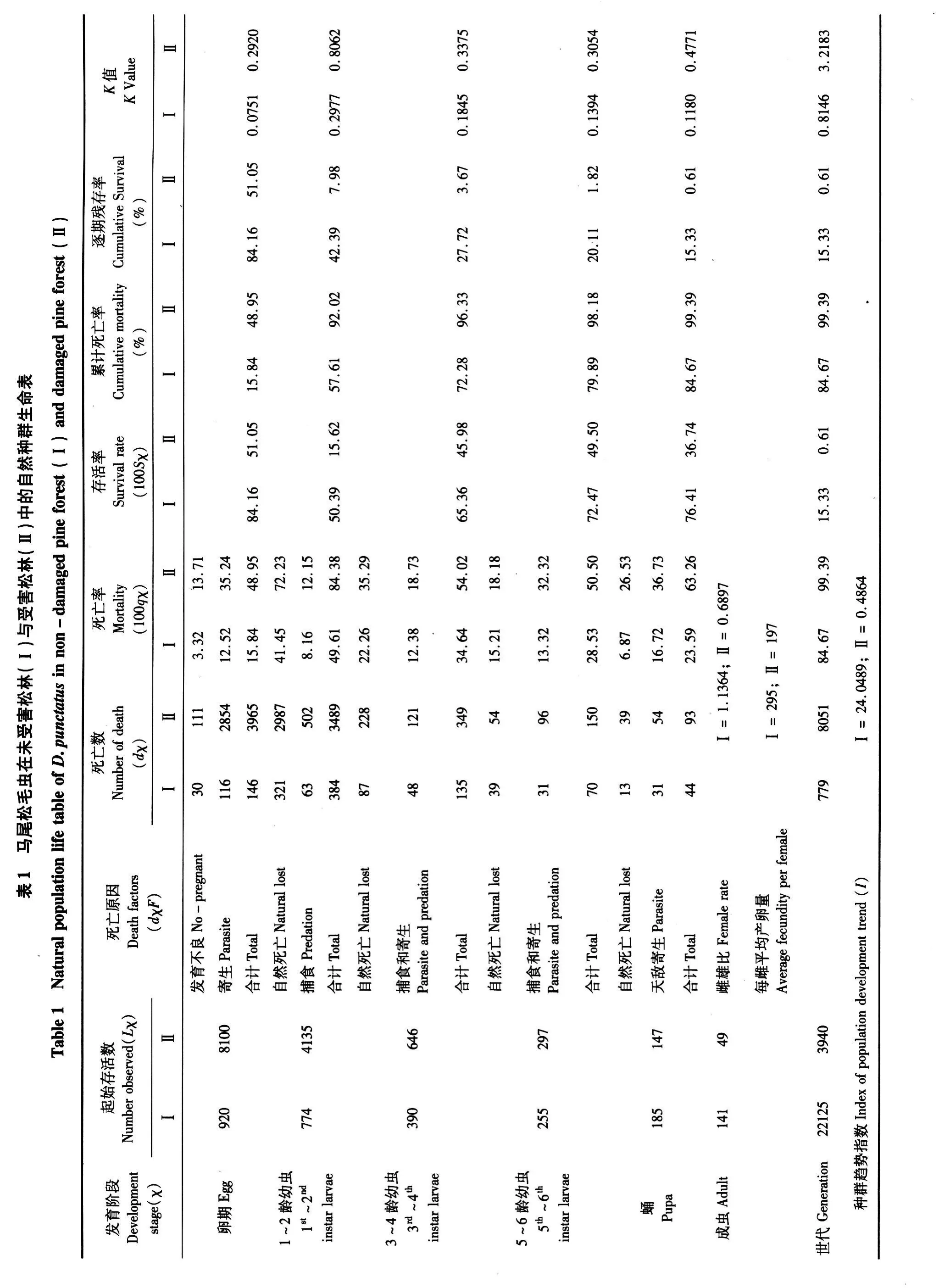
2.2 馬尾松毛蟲各發育階段消亡情況及關鍵因子分析
從自然種群生命表(表 1)可以看出,引起卵期死亡的主要原因為天敵的寄生作用。其中,馬尾松毛蟲在未受害松林中被天敵寄生導致的死亡率為12.52%,而在受害松林中達到 35.24%,表明在受害松林中,針葉的受害能加強對天敵的引誘功能,從而增強了天敵的寄生效能。
1~2齡幼蟲和 3~4齡幼蟲期死亡的主要原因為自然死亡,其次為天敵的捕食和寄生。其中,在受害松林中 1~2齡幼蟲的自然死亡率達 72.23%,3~4齡幼蟲的自然死亡率為 35.29%;分別為在未受害檢松林中同期幼蟲的自然死亡率的 1.74倍和1.58倍。5~6齡幼蟲期在被害松林中死亡的關鍵因子為天敵捕食和寄生,占該蟲期死亡率的 64%;而在未受害松林中死亡的主要原因仍為自然死亡,占死亡率的 70.88%。表明,馬尾松毛蟲幼蟲在被害松林中幼齡期由于自然因素導致蟲口的大量死亡且明顯高于在未受害松林中同期幼蟲的死亡率;同時,高齡幼蟲由于天敵的作用導致該蟲期死亡率要高于未受害松林中的幼蟲。
蛹期死亡的主要因子為天敵的寄生,其次為自然死亡。兩因子在受害松林中蛹期的死亡率分別為 37.63%和 26.53%;在未受害松林中的死亡率分別為16.72%和 6.87%。表明松毛蟲的蛹期在被害松林中受天敵的影響明顯高于未受害松林。
2.3 關鍵蟲期分析
根據馬尾松毛蟲各發育階段在兩類松林中死亡因素致死力(K值)的計算結果(表 1,圖 1)顯示,各因子在受害松林中的作用強度要明顯大于未受害松林,表現為在受害松林中的總 K值(3.2183)要遠遠大于未受害松林中的總 K值(0.8146)。然而,各因子作用的蟲期在不同類型的松林中表現不一。在受害松林中,馬尾松毛蟲的關鍵蟲期為 1~2齡幼蟲(K=0.8063),其次為蛹期(K=0.4771)和 3~4齡幼蟲期(K=0.3375);而在未受害松林中,馬尾松毛蟲的關鍵蟲期為 1~2齡幼蟲(K=0.2977),其次為 3~4齡幼蟲期(K=0.1845)和蛹期(K=0.1394)。
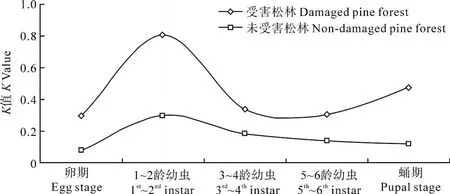
圖 1 馬尾松毛蟲數量變動關鍵因子分析Fig.1 Key factors analysis of D.punctatus population in non-damaged pine forest and damaged pine forest
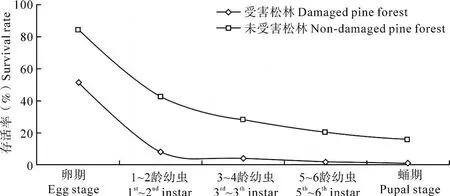
圖 2 在兩類松林中馬尾松毛蟲各發育階段的存活率Fig.2 Survival rate of D.punctatus population in non-damaged pine forest and damaged pine forest
2.4 存活曲線分析
根據表 1中馬尾松毛蟲各發育階段中的存活率繪制馬尾松毛蟲在不同類型松林中的存活曲線圖(圖 2),結果表明,兩類松林中馬尾松毛蟲自然種群的存活曲線均屬 Price所歸納的 A型(Price,1975)。在受害松林中,當代馬尾松毛蟲種群的殘存率只有 0.61%,而在未受害松林中有 15.33%的蟲體能羽化為成蟲。兩類松林中馬尾松毛蟲個體死亡主要發生在卵期和幼蟲期。其中,卵期在受害松林中的死亡率就達 48.95%,至 1~2齡幼蟲期只有 7.98%的個體存活。而在未受害松林中發育至 1~2齡幼蟲期還有 42.39%存活。表明受害松林對馬尾松毛蟲的生長發育不利而未受害的松林更有利于馬尾松毛蟲的生長。
3 結論與討論
國內外的研究表明,植物被害蟲危害后,會導致自身在外部形態、體內化學物質的變化,進而影響害蟲種群數量的增長(Haukioja and Neuvonen,1987;Haukioja,1990;Karban and Nhho,1997)。從組建的自然種群生命表中可以看出:馬尾松毛蟲的存活率和種群趨勢指數在未受害松林中要明顯高于受害松林,表明馬尾松毛蟲在未受害松林中取食易引起其種群數量的劇增,而在受害后的松林中繼續取食,其種群呈下降趨勢。自然死亡與天敵的捕食和寄生是導致馬尾松毛蟲種群死亡的主要原因,特別是在受害松林中,其總 K值要遠遠大于未受害松林中的總 K值,即兩因子在受害松林中的作用強度要明顯大于未受害松林。這些結果表明,受害松林中馬尾松毛蟲受自然因素以及天敵的捕食和寄生的控制較強,害蟲自然種群數量下降幅度較大,從而不利于馬尾松毛蟲種群的生長發育并對馬尾松毛蟲種群動態具有一定的自然調節作用。當然,這種抑制作用也與受害后松針內化學物質發生了變化導致馬尾松毛蟲的生長發育不良(王國紅等,2008)或增強了對天敵的引誘能力從而影響馬尾松毛蟲的種群增長(黃麗莉等,2006;王勇等,2008)。
自然種群消長規律即自然種群生命表的研究是害蟲管理的基本方法,對于馬尾松毛蟲自然種群消長規律的研究可以解決實際生產中的應用問題。松樹受害后,松樹本身化學成分發生變化,同時增強了對天敵的招引作用,使馬尾松毛蟲的自然種群呈下降趨勢,從而證明被害的松林自身具有一定的抵御蟲害的作用。雖然在被害松林中有松毛蟲的繼續危害,但在被害松林中的種群數量呈負增長趨勢而不致對松林造成很大的危害,故在生產中,可通過增加天敵因素,以生物防治為主導而避免使用化學藥劑。由于不同年份間氣候的變化有所不同,自然天敵種群數量發生消長亦有差異,因此,本文所建的生命表也只能反映大致的趨勢,今后應進一步應用多年的系列研究,從馬尾松毛蟲世代種群生命表中的完整數據進行系統分析,才能更全面、準確地評價被害松林對馬尾松毛蟲的種群影響機制。
References)
Agrawal AA,1998.Induced responses to herbivory and increased plant performance.Science,279(5354):1201-1202.
Haukioja E,1990.Induction of defence in trees.Annual Review of Entomology,36:25-42.
Haukioja E,Neuvonen S,1987.Insect population dynamics and induction of plant resistance:the testing of hypotheses.In:Barbosa P,Schultz TC,eds.Insect Outworks.San Diego:Academic,411-432.
Huang LL,Liu XP,Han RD,Wang GH,Sun JH,Ge F,2006.The effect of damaged and non-damaged pine needle on the tropism selection of parasitoid Trichogramma dendrolimi.Entomological Knowledge,43(2):215-219.[黃麗莉,劉興平,韓瑞東,王國紅,孫江華,戈峰,2006.松毛蟲赤眼蜂對被害與未被害馬尾松的趨性選擇.昆蟲知識,43(2):215-219]
Karban R,Baldwin IT,1997.Induced Responsesto Herbivory.Illinois,USA:University of Chicago Press,319.
Karban R,Nhho C,1993.Induced resistance and susceptibility to herbivory:Plant memory and altered plant development.Ecology,74(1):9-19.
Liu XP,Ge F,Chen CP,Wang GH,Li ZY,2003.Progress in induced resistance of pines.Scientia Silvae Sinicae,39(5):119-128.[劉興平,戈峰,陳春平,王國紅,李鎮宇,2003.我國松樹誘導抗蟲性研究進展.林業科學,39(5):119-128]
Lou YG,Cheng JA,1997.Induced plant resistance to phytophagous insects.Acta Entomologica Sinica,40(3):320-331.[婁永根,程家安,1997.植物的誘導抗蟲性.昆蟲學報,40(3):320-331]
Morris RF,Miller CA,1954.The development of life tables for the spruce budworm.Canadian Journal of Zoology,32:283-301.
Price PW,1975.Insect Ecology.Wiley,New York,514.
Wang GH,Liu XP,Ren Q,Hu YJ,2008.Content variation of secondary metabolites in damaged pinus massoniana needles and itseffect on the growth and development of Dendrolimus punctatus.Acta Agriculturae Universities Jianxiensis,30(4):581-585.[王國紅,劉興平,任琴,胡永建,2008.蟲害松針的次生物質變化及對馬尾松毛蟲生長發育的影響.江西農業大學學報,30(4):581-585]
Wang Y,Xiao TG,He Z,Ge F,2008.Effects of massion pinevolatiles on olfactory and parasitic behavior of Trichogramma dendrolimi.Entomological Knowledge,45(6):944-949.[王勇,肖鐵光,何忠,戈峰,2008.馬尾松樹針葉揮發性化學物質對松毛蟲赤眼蜂嗅覺及寄生行為的影響.昆蟲知識,45(6):944-949]
