不同基因型煙草品種接種赤星病菌后抗病蛋白的變化研究
2010-09-12 01:16:30曹祥煉宋彥君程海峰豆顯武楊鐵釗
浙江農業科學 2010年5期
關鍵詞:煙草
曹祥煉,宋彥君,程海峰,豆顯武,楊鐵釗
(1.湖北省襄樊市煙草公司 保康煙葉分公司,湖北 保康 441600;2.河南農業大學 煙草學院,河南 鄭州 450002)
煙草赤星病是由鏈格孢菌[Ternaria alternata(Fries)Keissler]引致的煙葉成熟后期重要的葉部真菌性病害,嚴重影響著煙草的產量和品質,制約著煙草生產的發展。赤星病在世界各煙草產區均有發生,是煙草生產上威脅最大的病害之一。赤星病具有潛育期短、流行速度快的特點,在環境條件有利于發病的情況下,短時間內即可造成大的流行,給煙葉生產帶來巨大損失[1]。煙草赤星病1892年首次在美國發現,曾幾次給世界煙葉生產帶來巨大的經濟損失[2]。1916年我國在北京附近首次發現,1964年在山東煙區流行,隨后全國各煙區有日益嚴重的趨勢。1989至1991年3年間,河南省平均每年因赤星病造成的損失達人民幣2 300萬元[3],成為毀滅性的病害。2000年8月初在美國的康乃狄克州和麻薩諸塞州,煙草赤星病大爆發,分別減產至少達75% 和89%[4]。
植物受到病原真菌侵染時,會產生一系列的主動防御反應。這些反應包括:合成植保素,感染區域細胞結構的加厚與增強,病程相關蛋白 (PR蛋白)的誘導和積累。這些反應還參與了防衛反應中的信號傳遞,使植物獲得系統性抗性[5-6]。
病程相關蛋白中研究較多的是幾丁質酶(Glu)和β-1,3-葡聚糖酶 (Cht)。很多植物病原真菌細胞壁的主要成分是幾丁質和葡聚糖,體外抑菌試驗表明,它們能抑制一些病原真菌孢子的萌發和病菌生長,因此幾丁質酶和葡聚糖酶一直被看作是植物抗真菌病害的潛在物質[5]。
1 材料與方法
1.1 試驗材料與種植
供試煙草品種為感赤星病品種長脖黃和高抗赤星病突變體凈葉黃,豫煙4號作感病對照,煙草品種來自河南省農業科學院。
試驗地點設在河南農業大學科教園區。種子、植煙土壤經過嚴格消毒,采用盆栽種植。3個品種分別于2008年5月10號移栽,重復3次,隨機區組排列,正常管理。
1.2 菌種與培養
孢子懸浮液。
1.3 接種與取樣
從煙草苗下部起選第3到第4片成熟葉片,噴霧接種,每個小區的各品種處理一致。接種后人工保溫30℃并保濕培養,分別于接種前0 h,接種后12,24,36和48 h取樣。以同生育期同部位成熟葉片噴施清水做空白對照。
田間采集到的赤星病菌強毒株在PDA培養基上培養7 d后,用無菌水洗下,配制成孢子濃度為106個·mL-1的孢子懸浮液。
1.4 測定方法
幾丁質酶活性的測定參照 Boller等[7]的方法。β-1,3-葡聚糖酶活性的測定參照史益敏[8]的方法。
2 結果與分析
2.1 幾丁質酶活性
幾丁質是許多真菌細胞壁、昆蟲體壁和甲殼類動物中的主要結構成分,但在高等植物中尚未發現有幾丁質的存在。當植物受到病原物侵染時體內會積累病程相關蛋白 (pathogenesis-relatedprotein,PR),已證明部分PR是幾丁質酶[10]。很多研究表明,幾丁質酶在參與寄主植物和病原菌之間相互作用有2個方面,即幾丁質酶有潛在的抗菌特性的直接作用和通過釋放幾丁質寡聚物而誘導植物產生抗性的間接作用[11]。現已有大量的試驗證實,植物幾丁質酶是可以被誘導的,大多數高等植物體內所含的幾丁質基因表達水平很低或不表達,而在受到如病原菌侵染、外源幾丁質、殼聚糖、水楊酸和乙烯等因子作用時均能被誘導表達[12]。
測定不同基因型的煙草品種接種赤星病菌后在正常生長條件下其葉片內幾丁質酶的基礎活性,結果如圖1所示:抗病品種凈葉黃幾丁質酶總活性的平均值為7個活力單位 (U),感病品種長脖黃為3個活力單位,而另一感病品種豫煙4號的幾丁質酶的基礎活性為4個活力單位左右。由此可見在正常生長條件下,抗病品種凈葉黃比感病品種長脖黃和豫煙4號體內幾丁質酶基礎活性高。

圖1 抗性煙草品種未接種幾丁質酶的變化
從圖2-4可以看出,無論是抗病品種還是感病品種,接種赤星病菌后幾丁質酶含量與其基礎含量相比都呈現明顯的上升趨勢。3個品種在接種赤星病菌后的前12 h幾丁質酶活性均上升較慢,凈葉黃在接種后12~24 h幾丁質酶含量達到10.06個活力單位,此后幾丁質酶活性仍一直上升,但升高速度減慢。感病品種長脖黃和豫煙4號幾丁質酶活性變化規律基本一致,在接種赤星病菌后48 h內酶活性呈上升趨勢。幾丁質酶潛在的抗菌特性的直接作用導致酶活性在接種前期劇烈上升,之后通過釋放幾丁質寡聚物而誘導植物產生抗性的間接作用,從而導致幾丁質酶活性的在接菌后的48 h內一直上升。從圖5可以看出,不同抗性煙草品種在接種赤星病菌后的前12 h,抗病品種凈葉黃幾丁質酶活性上升速度比感病品種長脖黃和豫煙4號要快,且在整個過程中凈葉黃的幾丁質酶活性都比長脖黃和豫煙4號要高,這可能是凈葉黃抗赤星病的原因。

圖2 凈葉黃接種后幾丁質酶的變化
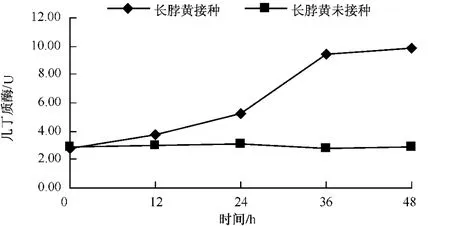
圖3 長脖黃接種后幾丁質酶的變化
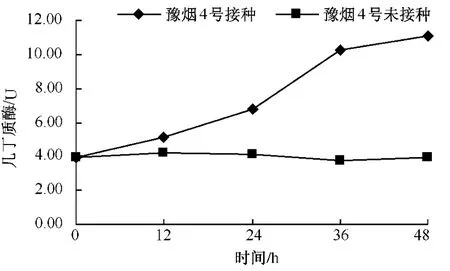
圖4 豫煙4號接種后幾丁質酶的變化
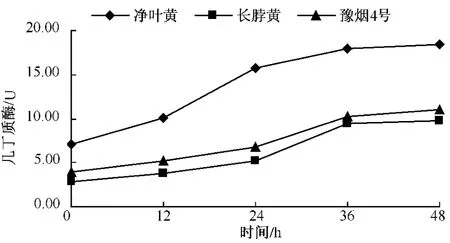
圖5 抗性煙草品種接種后幾丁質酶的變化
2.2 β-1,3-葡聚糖酶活性
測定不同基因型的煙草品種接種赤星病菌后,在正常生長條件下其葉片內β-1,3-葡聚糖酶的基礎活性,結果如圖6所示:不同基因型煙草品種β-1,3-葡聚糖酶活性變化與幾丁質酶基礎活性變化基本一致,都表現為長脖黃酶基礎活性最低,凈葉黃基礎活性最高,豫煙4號居中。抗病品種凈葉黃在正常生長條件其β-1,3-葡聚糖酶基礎活性的平均值為1 950個活力單位,感病品種長脖黃基礎活性為1 700個活力單位,而另一感病品種豫煙4號為1 850個活力單位上下。由此可見在正常生長條件下,抗性品種凈葉黃比感病品種長脖黃體和豫煙4號體內幾丁質酶的活性高。
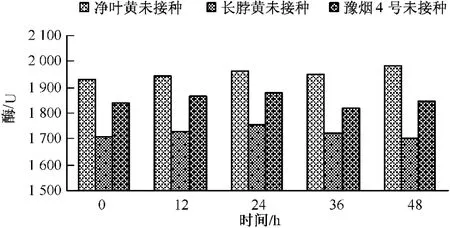
圖6 抗性煙草品種β-1,3-葡聚糖酶活性的變化
測定不同品種在接種赤星病菌后β-1,3-葡聚糖酶酶活性的變化,結果如圖7-9所示,無論是抗病品種還是感病品種,接種赤星病菌后β-1,3-葡聚糖酶含量與其基礎含量相比都呈現明顯的上升趨勢。凈葉黃在接菌后12 h內 β-1,3-葡聚糖酶達到2 272.4個活力單位,此后 β-1,3-葡聚糖酶活性仍一直上升,但升高速度減慢。這可能與葡聚糖在細胞壁內部外部均存在有關。感病品種長脖黃和豫煙4號β-1,3-葡聚糖酶活性變化規律基本一致,在接種赤星病菌48 h內酶活性呈上升趨勢。從圖10可以看出,不同抗性煙草品種在接種赤星病菌后的前12 h,抗病品種凈葉黃β-1,3-葡聚糖酶活性上升速度比感病品種長脖黃和豫煙4號要快,而感病品種長脖黃和豫煙4號在接種后12~24 h β-1,3-葡聚糖酶酶活性上升較快,但比抗病品種晚12 h。在整個過程中凈葉黃的β-1,3-葡聚糖酶活性都比長脖黃和豫煙4號要高,這可能是凈葉黃抗赤星病的原因。
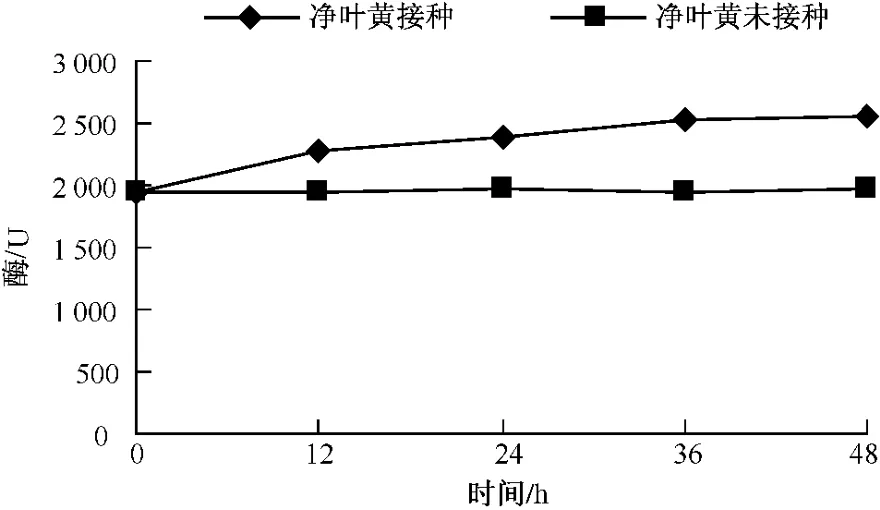
圖7 凈葉黃接種后β-1,3-葡聚糖酶活性的變化

圖8 長脖黃接種β-1,3-葡聚糖酶活性的變化

圖9 豫煙4號接種β-1,3-葡聚糖酶活性的變化
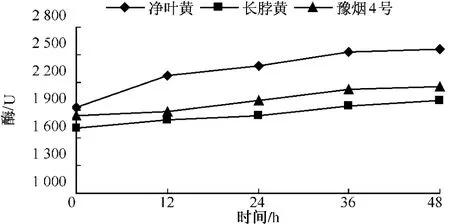
圖10 不同抗性品種β-1,3-葡聚糖酶活性的變化
2.3 幾丁質酶和β-1,3-葡聚糖酶活性變化趨勢
從圖11可以看出,凈葉黃在接菌后 β-1,3-葡聚糖酶變化比較平穩,幾丁質酶在接菌后12 h劇烈上升,24~48 h緩慢上升;感病品種長脖黃在接菌后,β-1,3-葡聚糖酶和幾丁質酶活性變化趨勢基本一致,在整個過程中一直呈上升趨勢,幾丁質酶在24~36 h劇烈上升,在36 h上升緩慢 (圖12);豫煙4號 β-1,3-葡聚糖酶活性在12~36 h上升明顯,幾丁質酶酶活性在24~36 h明顯增強(圖13)。從時間上看感病品種長脖黃和豫煙4號幾丁質酶酶活性劇烈上升比抗病品種凈葉黃晚12 h。3個品種在接菌后幾丁質酶和 β-1,3-葡聚糖酶酶活性均表現一定的相似性,幾丁質酶酶活性劇烈上升時間比β-1,3-葡聚糖酶晚。分析認為,可能與β-1,3-葡聚糖在病菌細胞壁內部外部都存在,而幾丁質主要存在與病菌的細胞壁內部有關。
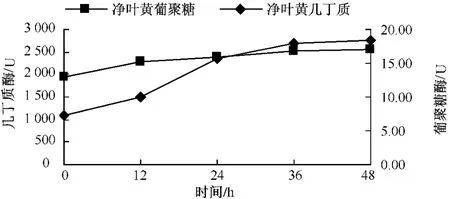
圖11 凈葉黃接種后幾丁質酶和β-1,3-葡聚糖酶酶活性的變化

圖12 長脖黃接種后幾丁質酶和β-1,3-葡聚糖酶酶活性的變化

圖13 豫煙4號接種后幾丁質酶和β-1,3-葡聚糖酶酶活性的變化
3 小結與討論
幾丁質和β-1,3-葡聚糖是真菌細胞壁的重要結構成分 ,在許多真菌的菌絲頂端 ,β-1,3-葡聚糖和幾丁質暴露在細胞壁表面 ,能夠直接受到β-1,3-葡聚糖酶和幾丁質酶的水解[14],這不僅使真菌菌絲生長點受到破壞 ,而且在水解過程中由真菌細胞壁釋放出來的寡糖能夠作為植物多種抗病反應的激發因子 ,誘導植物的全面防衛反應[14]。在正常生長條件下,抗病品種凈葉黃比感病品種長脖黃和豫煙4號體內幾丁質酶基礎活性高,接種赤星病菌后抗病品種凈葉黃幾丁質酶活性上升速度比感病品種長脖黃和豫煙4號要快。凈葉黃在接種后12 h幾丁質酶活性迅速升高,在之后的24 h幾丁質酶活性又一次劇烈上升。可能由于凈葉黃在赤星病菌接種初期,直接攻擊病原物真菌的細胞壁,使病原菌瓦解失去致病力,同時酶解后病原菌的細胞壁破碎物又可作為激發子再次刺激植物產生抗病防衛反應,導致酶活性又一次升高,從而達到抗病的目的。
在接種后的整個反應過程中長脖黃和豫煙4號的幾丁質酶活性與其對照相比都有不明顯的提高,但比凈葉黃的要低得多。分析認為赤星病菌侵染感病品種長脖黃和豫煙4號后,由于煙草體內被誘導的幾丁質酶的活性很低,抵抗和殺死病菌的能力差,是導致感病品種發病的原因之一。
無論是抗病品種還是感病品種,接種赤星病菌后β-1,3-葡聚糖酶含量與其基礎含量比都有明顯的上升。推斷由于接種后,隨著赤星病菌的入侵,煙草啟動體內有關的抗病防衛機制,相關抗病蛋白酶被激活。由于β-1,3-葡聚糖酶直接作用于赤星病菌的細胞壁,因此酶活性的升高對赤星病菌起到抑制作用。凈葉黃的β-1,3-葡聚糖酶的積累不管是在量上還是在時間上都比長脖黃和豫煙4號要快,對于病原菌的入侵凈葉黃反應較迅速,能及時抑制病原菌,阻止病害進一步擴展。從實驗結果可以看出,3個品種幾丁質酶積累時間與β-1,3-葡聚糖酶積累時間相比具有一定滯后性,可能與葡聚糖在真菌細胞壁內外都有,而幾丁質主要存在于細胞壁內部有關。
[1]Shew H D,Lucas G B.Compendium of tobacco disease[J].The American Phytopatho logical Society,1990,25:10-12.
[2]Stavely R,Echaplin J E,Gwynn G R.Registration of Bel 921 brown spot resistant flue-cured tobacco germplasm[J].Crop Science,1984,24:830-831.
[3]談文.煙草赤星病的發病規律及綜合治理[J].煙草科技,1993(2):45-48.
[4]Lamondia J A.Outbreak of brown spot of tobacco caused by alternaria alternata in connecticut and massachusetts[J].Plant Disease,2001,85(2):230.
[5]Kunze I,Nilsson C,Adler k,et al.Correct targeting of a vacudar tobacco chitinase in saccharomyces cerevisice posttranslational modifications are dependent on the rain[J].Biophys Acta,1998,1395(3):329.
[6]Lamb C J.Signals and transduction mechanismsfor activation of plant defeises against microbial attack[J]. Cell,1989,56:215.
[7]Boller T,Gehria A,Mauch F,et al.Chitinase in bean leaves:induction by ethylene,purification,properties, and possible function[J].Planta,1983,157:22 -31.
[8]史益敏.β-1,3-葡聚糖酶活性的測定[G]//中國科學院上海植物生理研究所.現代植物生理學實驗指南.北京:科學出版社,1999:128.
[9]Boller T. Chitinase in bean:induction by ethylene,purification,properties and possible function[J]. Plant,1983,157(1):22-31.
[10]趙蕾,汪天虹.幾丁質、殼聚糖在植物保護中的研究與應用進展[J].植物生理學通訊,1994,30(4):263-266.
[11]Benhamou N.菜豆內切幾丁質酶對立枯絲核菌的抗菌作用[J].微生物學雜志,1994(14):66-70.
[12]王維榮,裴真明,歐陽光察.幾種因子對黃瓜幼苗幾丁質酶的誘導作用[J].微生物學雜志,1994(14):66-70.
[13]Bonfante P, Perotto S, Teata B, etal. Ultrasturctural localization of cell surface sugar residues in ericoid mycowhizal fungi by gold lLabeled lectina[J].Protoplasma,2987,137:25-35.
[14]Selitrennikoff C P.Antifungal proteins[J].Apple Environ Microbiol,2001,67:2883-2894.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
