解脂耶氏酵母URA3基因的敲除
2010-08-27 11:13:46馮春利任清張蕾蕾李秀婷宋煥祿
食品與生物技術學報 2010年4期
馮春利, 任清, 張蕾蕾, 李秀婷, 宋煥祿
(北京工商大學化學與環境工程學院,北京 100048)
解脂耶氏酵母URA3基因的敲除
馮春利, 任清, 張蕾蕾, 李秀婷, 宋煥祿*
(北京工商大學化學與環境工程學院,北京 100048)
構建了營養缺陷型解脂耶氏酵母菌株,使之用于遺傳標記和高產香味物質γ-癸內酯。作者利用基因同源重組的方法敲除掉尿嘧啶合成酶關鍵基因URA3基因,用尿嘧啶營養缺陷型培養基(SD-URA)添加一定濃度的5-氟乳清酸(5-FOA)和尿嘧啶篩選獲得轉化子。實驗表明:尿嘧啶營養缺陷型菌株在含有5-FOA和尿嘧啶的培養基上生長而野生型菌株不生長,從而建立了一種快速獲得營養缺陷型解脂耶氏菌株的方法。
解脂耶氏酵母菌;URA3;基因敲除
解脂耶氏酵母于1942年首次被分離得到,先后被命名為Candida lipolytica、Endomycopsis lipolytica、S accharomycopsis lipolytica,最終定名為Yarrowia lipolytica(解脂耶氏酵母)[1]。解脂耶氏酵母是非常規酵母中的一種[2],非常適應疏水環境,可以代謝不飽和羥基酸,用于香味物質γ-癸內酯的制備。在眾多的可以生產γ-癸內酯的菌株中,酵母Yarrowia lipolytica的發酵能力是較強的[3]。
在Yarrowia lipolytica中,URA3[4-5]基因能夠編碼乳清苷酸脫羧酶基因,該酶是尿嘧啶合成的關鍵酶。該酶又可催化5-氟乳清酸(5-FOA)[6]轉化為有毒物質。作者通過對URA3基因敲除,使5-氟乳清酸(5-FOA)無法形成有毒物質5-氟尿嘧啶核苷酸,從而對5-FOA具有抗性,其嘧啶核苷酸營養則可以向培養基加入尿嘧啶通過補救途徑給與補充;而5-FOA可以抑制野生型解酯酵母菌生長。因此5-FOA可用于尿嘧啶營養缺陷型突變體的篩選[7-8]。
作者構建的營養缺陷型解脂酵母菌株,一方面可以作為遺傳標記,應用于遺傳工程和菌種改良。另一方面由于解酯酵母菌一般用于香味物質γ-癸內酯的制備。蘇暢等[9]曾對7株菌產生γ-癸內酯的能力進行了研究,結果發現Yarrowia lipolyticaAS2.1405產生γ-癸內酯的能力最強,產率為1%。作者所在實驗室所用的菌株為Yarrowia lipolyticaAS2.1405。在不含有尿嘧啶的培養基中,尿嘧啶缺陷型的Yarrowia lipolytica菌株進行生物轉化蓖麻油酸甲酯,使蓖麻油酸的一個旁路代謝途徑和細胞生長受到抑制,從而轉回內酯的合成途徑[10],由此可以提高香味物質γ-癸內酯產量,具有非常重要的理論價值和應用前景。
1 材料與方法
1.1 菌種及質粒
解脂耶氏酵母Y.lipolyticaAs 2.1405:購自中國模式菌種保藏中心,是一株模式菌株,為非常規酵母,用于香味物質γ-癸內酯的制備;大腸桿菌DH5α:作者所在實驗室保藏;質粒pBluscriptks +:作者所在試驗室保藏,用于在大腸桿菌中載體制備,具有氨芐青霉素抗性,可用于大腸桿菌的藍白篩選。
1.2 培養基
YEPD液體培養基(組分g/L):葡萄糖1,蛋白胨1,酵母浸膏0.5;p H值自然。112℃滅菌20 min。
LB液體培養基(組分g/L):蛋白胨1,酵母浸膏0.5,NaCl 1。121℃滅菌20 min。
LB固體培養基:LB液體培養基中加1.5 g/dL的瓊脂。
GENMED酵母菌尿嘧啶合成缺陷型(SDURA)培養基:含有除尿嘧啶外所有其它氨基酸和核苷酸基本營養元素,用于尿嘧啶營養缺陷型的篩選,購自上海百事生物科技有限公司。
1.3 酶及主要試劑
蝸牛酶、Taq DNA Polymerase聚合酶、T4 DNA Ligase連接酶等:均購于北京天根生化科技有限公司。Trans DNA MarkerⅢ:購于北京全式金生物技術有限公司;限制性內切酶:SalⅠ、EcoRⅠ、PstⅠ、HindⅢ等酶:購于寶生物工程(大連)有限公司;小牛胸腺DNA,5-FOA,PCR擴增所用引物等:均購自上海生工有限公司;其他化學試劑:購自北京拜爾迪生物技術有限公司。
1.4 引物設計
利用NCBI的Entrez檢索系統,查知該酵母菌已經注冊過的URA3的基因,獲得啟動子和終止子序列;根據已知序列,用Primer premier5.0軟件設計上游引物和下游引物。
URA3上游引物:5’-CAA GGTACCGCTATCACATCACGCTCTCA-3’,下劃線部分為KpnⅠ酶切位點。
URA3下游引物:5’-ATCGAATTCTCATGA TTTCAAACACG-3’,下劃線部分為EcoRⅠ酶切位點。
1.5 主要儀器及設備
Alpha自動凝膠成像儀:Alpha Inotech公司制造;DYY-6C電泳儀:北京六一儀器廠制造;XP梯度PCR儀:杭州博日科技有限公司制造。
1.6 方法
1.6.1 酵母基因組的提取 將保存在斜面培養基上的解脂酵母菌接種于2 mL液體YEPD培養基中,28℃振蕩過夜(200 r/min),提取酵母基因組[11],0.8 g/dL瓊脂糖凝膠電泳檢測。
1.6.2 目的基因的PCR擴增 利用PCR方法擴增Yarrowia lipolytica尿嘧啶合成酶關鍵基因URA3。
PCR反應條件為:95℃預變性5 min;94℃變性4 s,55℃退火45 s,72℃延伸2 min 45 s,循環30次;72℃最后延伸10 min,4℃保存。
1.6.3 基因敲除組件的構建
1)質粒pBluscript ks+和PCR擴增產物URA3基因的酶切:PCR產物酶切體系20μL為10 ×M Buffer 2μL、PCR產物10μL、去離子水6.6 μL、EcoRⅠ0.7μL、KpnⅠ0.7μL;質粒pBluscript ks+酶切體系20μL為10×M Buffer 2μL、質粒DNA 2μL、去離子水14.4μL、EcoRⅠ0.8 μL、KpnⅠ0.8μL;37℃保溫5 h,然后電泳。將質粒和PCR產物酶切體系用天根瓊脂糖凝膠回收試劑盒回收,然后放入-20℃保存備用。
2)重組質粒m13-URA3的酶連:在1.5 mL離心管中加入以下20μL體系。目的DNA片段12 μL、載體DNA 3μL、10×Ligase Buffer 2μL、T4 DNA Ligase 2μL、去離子水1μL。調節保溫杯為16℃,連接過夜。
3)重組質粒的轉化:將酶連產物轉化大腸桿菌E.coli DH5α感受態細胞,方法見文獻[11,12]。
4)重組質粒快速提取:用無菌牙簽挑菌法挑取含有氨芐青霉素LB平板上的白色菌落,接種于含Amp 50μg/mL的2 mL LB培養基中,37℃振蕩過夜。取出大腸桿菌至1.5 mL離心管中,離心加25μL TE緩沖液再加入25μL Tris飽和酚-氯仿(體積比1∶1),激烈振蕩混勻后12 000 r/min離心5 min,用1 g/dL瓊脂糖凝膠電泳驗證。
5)重組質粒的大量提取與鑒定:提取小量快速提取后比原始質粒大的重組質粒,并進行單酶切和雙酶切驗證。
6)URA3基因敲除組件的構建:將已經構建的重組質粒m13-URA3進行SalⅠ單酶切,由于在URA3基因的CDS區兩側有相同的SalⅠ酶切位點,可以將URA3基因的CDS區切除,從而進一步16℃連接過夜。轉化大腸桿菌后大量提取已經去掉URA3的CDS區的基因敲除質粒。
1.6.4 將URA3基因敲除組件轉化酵母 制備酵母菌感受態細胞,利用醋酸鋰方法[13-14]介導URA3基因敲除組件轉化酵母感受態細胞,轉化后的酵母菌涂布于含有5-FOA的GENMED酵母菌尿嘧啶合成缺陷型(SD-URA)培養基上,28℃培養3~4 d使轉化子形成單菌落。同時做一組未進行轉化質粒的酵母菌對照。
1.6.5 尿嘧啶營養缺陷型菌株的篩選和分子生物學鑒定 從合成缺陷型(SD-URA)培養基上挑取單菌落進行饑餓培養至菌體濃度不再增長,將其均勻涂布于加有尿嘧啶的完全培養基和不添加尿嘧啶的基本培養基上做對比。經多次復篩和分離純化獲得遺傳性狀穩定的尿嘧啶營養缺陷型菌株進行保存。并將這些菌株進行提取基因組后,利用原URA3基因上下游引物進行PCR擴增,進行分子生物學鑒定。
2 結果與討論
2
.1 目的基因的PCR擴增
利用設計好的引物1和引物2,對已經提取的酵母基因組進行URA3基因的PCR擴增,并獲得與NCBI上已經公布的該基因大小相符,經天根生物技術公司基因測序,為所需要的目的基因,長度為2.56 kb,見圖1。
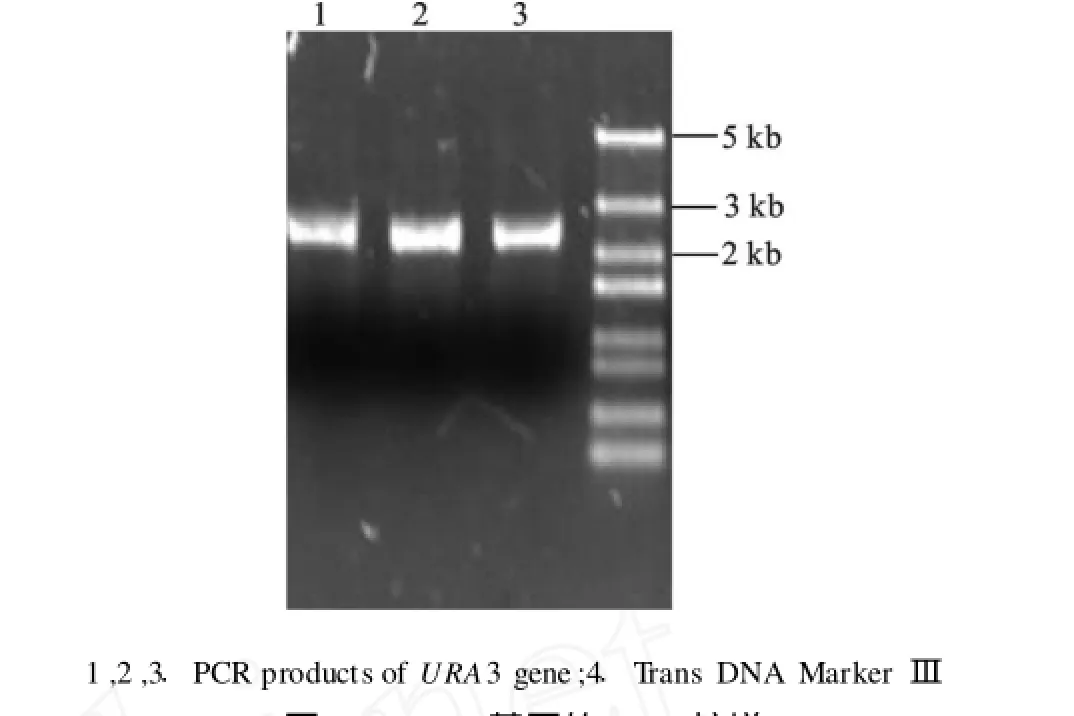
圖1 URA3基因的PCR擴增Fig.1 PCRamplification ofURA3 gene
2.2 構建URA3基因敲除組件
首先將大小為2.56 kb左右的URA3基因連到pBluscript ks+載體上,見圖2。由于在URA3基因上有兩個SalI酶切位點,將5.5 kb的m13-URA3重組質粒進行Sal I酶切,可以將URA3的CDS區切除,凝膠回收3.8 kbp左右片段,然后酶連獲得去除CDS區的重組敲除載體。其片段大小為3.8 kb左右。構建的質粒酶切驗證見圖3。左側為pBluscript ks+載體和URA3基因連接后的酶切圖。1道為m13-URA3重組質粒載體,2道為用SalⅠ酶切后將URA3基因的CDS區切除,大小分別為3.8 kb和1.7 kb。將3.8 kb片段凝膠回收,然后用T4連接酶于16℃連接構成敲除組件。
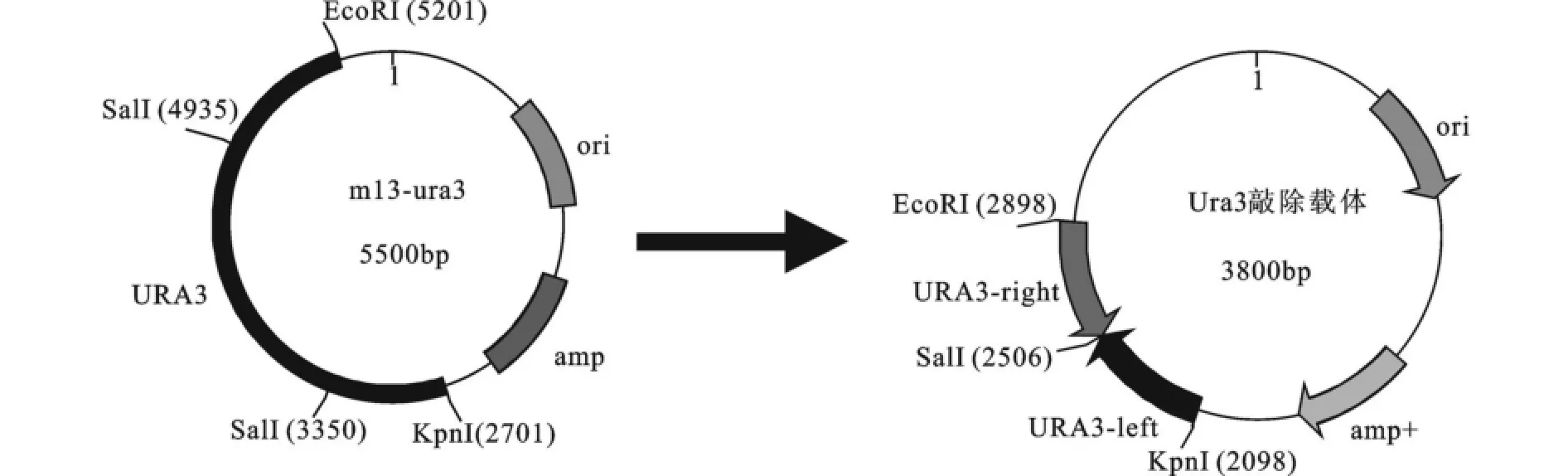
圖2 URA3基因敲除組件的構建過程Fig.2 Construction ofURA3 gene disrupt cassette
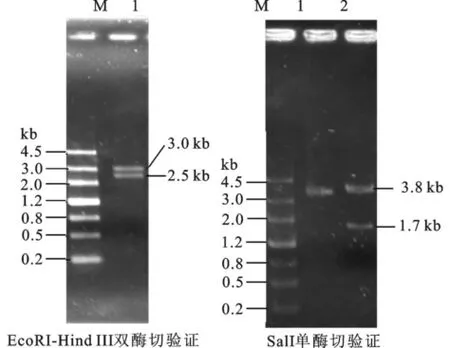
圖3 重組質粒m13-URA3質粒的單雙酶切驗證Fig.3 Recombinantm13-URA3 plasmid digestion validation
2.3 利用基因敲除組件轉化酵母
將獲得的基因敲除組件用PstⅠ進行單酶切,將URA3基因敲除組件切成線型,轉化酵母。此敲除組件與酵母體內URA3基因左側有454 bp,右側有395 bp的同源片段,因此可以發生同源重組。從而替換掉酵母內部的URA3基因,用含有一定濃度5-FOA和尿嘧啶的篩選培養基篩選敲除掉URA3基因的營養缺陷型轉化子。表1、2為進行初步篩選。

表1 敲除組生長情況Tab.1 G rowth of the disrupted strain

表2 野生型(未敲除組)生長情況Tab.2 G rowth of the wild type strain
將初篩獲得的轉化子挑取菌株接入不含尿嘧啶的液體培養基和固體培養基上,均未生長,同時做了對照組含有尿嘧啶和5-FOA為1 mg/mL的培養基,結果轉化子生長,說明初篩獲得的營養缺陷型轉化子是正確的。
2.4 尿嘧啶營養缺陷型菌株的進一步驗證和分子生物學鑒定
提取營養缺陷型酵母基因組和野生型酵母基因組,用URA3基因上下游引物對其分別進行PCR擴增,并且在含有尿嘧啶20μg/mL和5-FOA為1 mg/mL的(SD-URA)固體培養基上劃線培養。
由圖4可知,營養缺陷型菌株進行URA3基因PCR擴增得到849 bp片段,而野生型菌株PCR擴增片段仍為2 560 bp。圖5左側為野生型菌株,未生長,而右側為獲得的營養缺陷型菌株。由此雙重驗證了基因敲除成功,構建出了尿嘧啶營養缺陷型菌株。
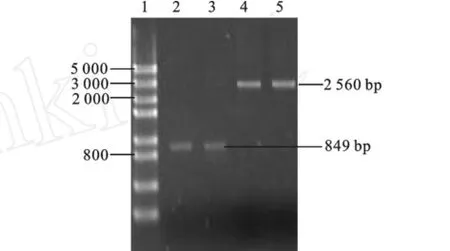
圖4 基因缺陷型的分子生物學驗證Fig.4 G enetic defects in molecular biology-based authentication
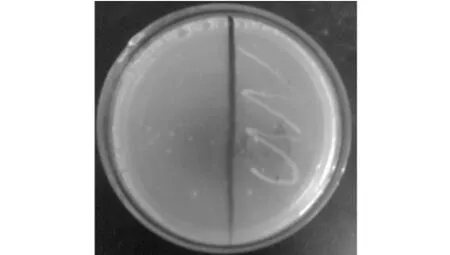
圖5 尿嘧啶營養缺陷型菌株的鑒定Fig.5 Identification of uracil auxotrophic strain
3 結 語
構建了2個重組質粒并且均經過了單雙酶切的驗證,然后通過同源重組的方法轉化酵母獲得營養缺陷型轉化子。在敲除掉URA3基因后,可用其作為篩選標記,進行其他功能基因的敲除研究。通過基因敲除技術得到的尿嘧啶營養缺陷型菌株,在不含有尿嘧啶的發酵培養基中,能夠抑制蓖麻油酸的一個旁路代謝途徑和細胞生長,使其轉回內酯的合成途徑,從而提高香味物質γ-癸內酯產量,因此在下一步生產實踐中,還可以進行培養基優化,進而使γ-癸內酯產量更加提高,進行香味物質制備。
[1]Barth G,Gaillardin C.Non-conventional Yeasts in Biotechnology[M].Berlin:Springer-Verlag,1996:313-388.
[2]Madzak Catherine,Gaillardin Claude,Beckerich Jean-Marie.Heterologous protein expression and secretion in the non-conventional yeastYarrowia lipolytica:a review[J].Journal of Biotechnology,2004,109:63-81.
[3]Wache Y,Aguedo M,Nicaud J M.Catabolism of hydroxyacids and biotechnological production of lactones byYarrowia lipolytica[J].Applied Microbiology and Biotechnology,2003,61:393-404.
[4]LI Wen-li,LIU Gang,TAN Hua-rong.Disruption of sabR affects nikkomycin biosynthesis and morphogenesis inStreptomyces ansochromogenes[J].Biotechnology Letters,2003,25:1491-1497.
[5]Kim J G,Choi YD,Chang YJ,et al.Genetic transformation ofMonascus purpureusDSM1379[J].Biotechnology Letters,2003,25:1509-1514.
[6]白玉杰.酵母雙雜交系統研究及其應用[Z].西安:第四軍醫大學生物化學及分子生物學教研室,2002.
[7]Boeke J D,LaCroute F,Fink G R.A positive selection for mutants lacking orotidine-5’-phosphate decarboxylase activity in yeast:5-fluoro-orotic acid resistance[J].Mol G en G enet,1984,197:345-346.
[8]Peck R F,Krebs M P.Homologous gene knockout in the archaeonHalobacterium salinarumwithURA3 as a counterselectable marker[J].Molecular Microbiology,2000,35(3):667-676.
[9]蘇暢,杜毅,陳洪.微生物發酵法生產γ-癸內酯[J].食品工業科技,2004,10:118-189.
SU Chang,DU Yi,CHEN Hong.The production of gamma-decalactone by microbial fermentation[J].Science and Technology of Food Industry,2004,10:118-189.(in Chinese)
[10]Pagot Y,Endrizzi A,Nicard J M.Utilization of an auxotrophic strain of the yeastYarrowia lipolyticato improveγ-decalactone production yields[J].Letters in Applied Microbiology,1997,25:113-116.
[11]J.薩姆布魯克,D.W.拉塞爾.分子克隆實驗指南[M].北京:科學出版社,2002:385-455.
[12]李欣,楊坤寧,劉志勇,等.電轉化法制備高轉化效率的E.coli感受態細胞研究[J].食品與生物技術學報,2007,11(6): 48-51.
LI Xing,YANG Kun-ning,LIU Zhi-yong,et al.Study on preparation of high transformation efficiency electrocompetentEscherichia coli[J].Journal of Food Science and Biotechnology,2007,11(6):48-51.(in Chinese)
[13]Gietz R D,Woods R A.Transformation of yeast by the lithium acetate/single-stranded carrier DNA/polyethylene glycol method[J].Methods Enzymol,2002,350:87-96.
[14]Maitreya T,Takegawa K.A simple and efficient procedure of transformation ofSchizosaccharomyces pombe[J].Yeast, 2004,21(8):613-617.
(責任編輯:李春麗)
The Disruption ofURA3 Gene ofYarrowia lipolytica
FEN G Chun-li, REN Qing, ZHANG Lei-lei, LI Xiu-ting, SONG Huan-lu*
(College of Chemical&Environmental Engineering,Beijing Technology&Business University,Beijing 100048,China)
In order to get high-yieldγ-decalactone and genetic markers,an auxotrophicYarrowia lipolyticastrain was isolated by genetic modification.In this study,URA3,a key gene for uracil synthetase ofYarrowialipolytica,was disrupted using the method ofgene homologous recombination,then transformants were screened by uracil auxotrophic medium(SD-URA)with the combination of 5-FOA and uracil.The results showed that the uracil auxotrophic strain was able to grow at the SD-URA medium containing 5-FOA and uracil while the wild-type strains do notgrown.Therefore,an easy and efficientmethod for obtaining auxotrophicYarrowia lipolyticastrain was established in this study.
Yarrowia lipolytica,URA3,gene disruption
Q 819
:A
1673-1689(2010)04-0624-05
2009-09-01
北京市教委科技計劃重點項目(KZ200910011001)。
*通信作者:宋煥祿(1961-),男,山東煙臺人,工學博士,教授,碩士生導師,主要從事食品風味化學及農產品深加工方面的研究。Email:songhl@th.btbu.edu.cn
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
今日農業(2021年11期)2021-08-13 08:53:34
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學一年級版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
海峽姐妹(2016年5期)2016-02-27 15:20:20
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
