幾種腫瘤特異性啟動子在人肝癌細胞系中的活性比較*
2010-06-15 01:44:00黃愛龍
重慶醫學 2010年18期
關鍵詞:肝癌
況 舸,唐 霓,陶 鵬,吳 瑩,張 君,黃愛龍△
(重慶醫科大學:1.藥學院藥理教研室 400016;2.教育部感染性疾病分子生物學重點實驗室,病毒性肝炎研究所 400016;3.附屬第二醫院感染科 400010)
原發性肝癌是一種常見的惡性腫瘤,適于手術切除的病例很少,常規的手術、化療、放療療效較差。肝癌基因治療是研究的熱點,多種基因治療手段已經被證實具有良好的腫瘤殺傷作用,但由于目前采用的基因治療載體缺乏腫瘤靶向性,因此限制了基因治療方法的臨床應用。
利用腫瘤特異性啟動子調控治療基因表達是解決基因治療靶向性的方法之一,目前研究最多的是甲胎蛋白(AFP)啟動子在肝癌的基因治療中的應用。多項研究已經證實AFP基因的調控區能夠在肝癌細胞內特異性激活外源基因的轉錄,并在肝癌的靶向性基因治療中取得一定療效[1-2]。但AFP啟動子存在轉錄活性低、陽性率低的弱點。因此,肝癌的靶向性基因治療需要尋找更加廣譜、有效的啟動子來調控目的基因的表達。
端粒酶的激活是細胞發生永生化及癌變過程中極為重要的步驟。人端粒酶逆轉錄酶(human telomerase reverse transcriptase,hTERT)僅在端粒酶陽性的腫瘤細胞和永生化細胞中表達,是端粒酶的催化亞基,決定了端粒酶的活性[3]。在所有肝細胞癌(hepatocellular carcinoma,HCC)中均有hT ERT表達,其中74%為強表達[4]。hTERT基因上游調控區具有強啟動子活性,且僅表現于端粒酶陽性的永生化及癌細胞系,提示hTERT基因啟動子具有腫瘤特異性的轉錄活性[5]。
survivin(SUR)是一類僅在腫瘤細胞中表達的凋亡抑制因子(inhibitor of apoptosis,IAP),在終末分化的成人組織不表達[6]。在12種常見的肝癌細胞株中SUR mRNA均有表達,SUR蛋白在70%肝癌中呈強陽性,其表達在肝癌組織與癌旁肝組織間存在顯著性差異[7-9]。因此,SUR啟動子被認為是一種肝癌特異性的啟動子,能夠實現治療基因在肝癌中的選擇性轉錄,避免損傷正常肝組織。
作者用PCR方法從人類基因組中克隆了hTERT、SUR、AFP基因 ATG上游的啟動子序列,分別構建了 hTERT、SUR、AFP驅動的含有報告基因蟲熒光素酶(luciferase,Luc)的真核表達載體,通過測定在人HCC細胞系和正常原代培養細胞中的Luc的表達,評價并比較hTERT、SUR、AFP啟動子的轉錄活性,為進一步應用于肝癌靶向性基因治療打下基礎。
1 材料與方法
1.1 材料
1.1.1 質粒和細胞 pGL3-Basic和 pRL-tk購自 Promega公司,前者為無啟動子的含有Luc基因的質粒,后者為含有單純皰疹病毒胸苷激酶(HSV-TK)啟動子控制的海洋腔腸熒光素酶基因(renilla luciferase,RL)的質粒。肝癌細胞系HepG2、QGY7701和SMMC7721均由本研究所保存。
1.1.2 主要試劑 T4DNA連接酶、各種限制性內切酶、雙熒光檢測系統Dual-Luciferase Reporter Assay System購自Promega公司,脂質體轉染試劑Lipofectamine2000購于Invitrogen公司。
1.2 方法
1.2.1 HepG2細胞基因組DNA的提取 單層培養的HepG2細胞長至80%融合,PBS洗3次后用細胞刮子刮下收集于1.5 mL的EP管中。按DNA提取試劑盒操作說明制備基因組DNA。
1.2.2 AFP、SUR、hTERT巨細胞病毒和巨細胞病毒(CM V)啟動子引物設計 根據GenBank中SUR基因和AFP基因序列以及pEGFP質粒中CMV啟動子序列設計引物并分別在各上下游引物5′端引入BglⅡ和HindⅢ酶切位點。hTERT啟動子的引物序列:上游5′-CGC AGA TCT CAG CTG CGC TGT CGG GGC CAG G-3′;下游 5′-CGC AAG CT T GCT GCC TGA AAC TCG CGC CGC GA-3′。SU R啟動子的引物序列:上游 5′-CGC AGA TCT ATT ACA GGC ATG CAC CAC-3′;下游 5′-GCG AAG CTT AAT CTG GCG GTT AA T GGC-3′。AFP啟動子的引物序列:上游 5′-GCG AGA TCT TCT GCA ACT TAG GGA CAA-3′;下游 5′-GCG AAG CT T GT T ATT GGC AGT GGT GGA-3′。 CM V 啟動子引物序列:上游 5′-GCG AGA TCT AA T CAA TTA CGG GGT CAT-3′;下游 5′-GCG AAG CT T GGA TCT GAC GGT TCA CTA-3′。
1.2.3 AFP、SUR、hTERT和 CMV啟動子片段的PCR擴增采用50 μ L 體系。以 1 μ g DNA 為模板;分別加入 Taq酶1.5 u;上下游引物各50 pmol;dNTPs 10 nmol。以 HepG2基因組DNA為模板擴增 AFP、SUR、hTERT啟動子片段,以質粒pEGFP為模板擴增 CMV啟動子。AFP、SUR、hT ERT啟動子片段擴增采用熱啟動法。AFP啟動子擴增條件為:95℃,4 min,加入Taq酶。94℃,45 s;60℃,50 s;72℃,1 min。25個循環后,72℃延伸10 min。PCR產物為330 bp。hT ERT啟動子擴增條件為:95℃,4 min,加入 Taq酶。94℃,45 s;72℃,50 s;72℃,60 s。25個循環后,72℃延伸10 min。PCR產物為280bp。SUR啟動子擴增條件為:95℃,4 min,加入Taq酶。94℃,45 s;62.5℃,50 s;72℃,60 s。25個循環后,72℃延伸10 min。PCR產物為414 bp。CM V啟動子擴增采用普通PCR法,擴增條件為:94℃,5 min。94℃,45 s;56℃,50 s;72℃,60 s。25個循環后,72℃延伸10 min。PCR產物為594 bp。
1.2.4 報告基因真核表達載體 pGL3-AFP、pGL3-SUR、pGL3-hTERT和 pGL3-CMV的構建 PCR產物經回收和BglⅡ-HindⅢ雙酶切后,克隆至pGL3-Basic載體的相應位點。重組質粒經XhoⅠ/HindⅢ雙酶切鑒定后送上海博亞公司測序。測序正確的克隆分別命名為pGL3-AFP和pGL3-SUR、pGL3-hTERT和pGL3-CMV。構建過程見圖1。
1.2.5 細胞轉染和熒光素酶報告基因檢測 以pGL3-Basic為陰性對照,以 pGL3-CMV為陽性對照,將 pGL3-Basic、pGL3-SUR、pGL3-AFP、pGL3-hT ERT 和 pGL3-CM V 各 1ug分別與 pRL-tk 0.1 μ g 共轉染 HepG2、SM MC7721、QGY7701細胞和原代培養的成纖維細胞。HepG2、QGY7701和SMMC7721均培養于含10%熱滅活小牛血清的RPM I1640培養基,原代成纖維細胞培養于含10%胎牛血清的DMEM培養基。于37 ℃,5%CO2恒溫培養箱培養。將 HepG2、QGY7701、SMMC7721和原代成纖維細胞按0.6×105/孔種于24孔板中,用無抗生素的培養液培養24 h后,換無血清培養液,用脂質體(Lipofectamin2000)轉染,每種質粒轉染 4孔。轉染6 h后換掉轉染培養液,用含10%小牛血清的RPMI1640培養液繼續培養。24 h后,用裂解液裂解轉染細胞,測定Luc和RL活性。后者的活性反映每孔細胞的轉染效率。各啟動子的相對活性以Luc/RL的比值來比較的,同一轉染實驗的4孔所得Luc/RL的比值求平均值和標準差,并據此作圖。
2 結 果
2.1 AFP、hTERT、SUR和CMV 基因啟動子的擴增 以HepG2基因組DNA為模板進行PCR擴增。所獲PCR產物進行1%瓊脂糖凝膠電泳分析(圖1)。結果表明,PCR擴增產物分別位于約330、280、465 bp及約595 bp處。

圖1 AFP、hTERT、SUR、CM V啟動子 PCR產物瓊脂糖凝膠電泳
2.2 重組質粒的鑒定 酶切鑒定結果見圖2、3。所有質粒均送上海博亞公司測序。測序結果證實此4個啟動子序列與目的啟動子序列完全一致,無堿基突變。
2.3 細胞轉染和熒光素酶報告基因的檢測 檢測結果見表1。
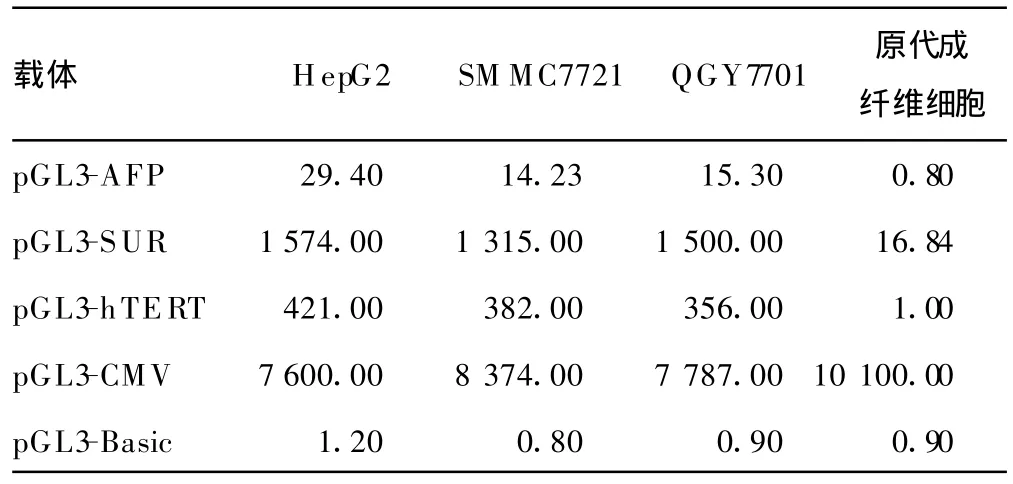
表1 AFP、hTERT、SUR和CMV啟動子的相對活性(%)

圖2 BglⅡ+HindⅢ雙酶切鑒定重組質粒pGL3-AFP、pGL3-SUR、pGL3-CMV

圖3 BglⅡ+HindIII雙酶切鑒定重組質粒 pGL3-hTERT
結果顯示:AFP、hTERT和SU R的啟動子均是有腫瘤特異性轉錄活性。CM V啟動子的轉錄活性沒有腫瘤特異性。在3種具有腫瘤特異性轉錄特性的啟動子中,SUR啟動子活性最高,hTERT啟動子次之,AFP啟動子活性最低(圖4)。在HepG2細胞中,SUR啟動子活性是AFP的53倍,是hT ERT啟動子活性的3.7倍,相當于CMV的21%;在SM MC7721細胞中,SUR啟動子活性是AFP的92倍,是hT ERT啟動子活性的3.4倍,約占CMV的 16%;在QGY7701細胞中SU R啟動子活性是AFP的98倍,是hTERT啟動子活性的4.2倍,達到CMV的19%(表1)。而hTERT啟動子在三種肝癌細胞系中的活性分別為AFP啟動子的14倍、30倍和24倍,相當于CM V啟動子活性的5.5%、4.6%、4.6%。SUR和hTERT啟動子的活性較最常用的肝癌特異性啟動子AFP啟動子有顯著增加,即使在高表達AFP的HepG2細胞中,其啟動活性也比AFP啟動子活性高數十倍。以上結果提示SUR和hTERT啟動子在肝癌細胞中均能夠高度激活外源基因表達,在高表達AFP的肝癌細胞中,其啟動子活性也優于AFP啟動子,見圖4。

圖4 AFP、hTERT和 SUR啟動子在 HCC細胞中的轉錄活性比較
3 討 論
基因治療能否成功應用于臨床實踐最為關鍵的問題是如何提高治療基因的靶向性和表達效率,以保證治療的安全性和有效性。利用腫瘤特異性啟動子調控目的基因在腫瘤細胞中的表達是實現靶向性基因治療的重要手段。目前肝癌的靶向基因治療中應用最多的是AFP啟動子。但AFP啟動子僅在高表達AFP的肝癌細胞中具有一定的治療效果;對低表達AFP的肝癌無效[1-2]。而在人類肝癌中,高表達AFP的僅占40%左右。因此,對于肝癌的基因治療需要尋找更加廣譜、有效的啟動子來調控目的基因的表達。
端粒酶是一種新的廣譜的腫瘤標志物,約90%的腫瘤中端粒酶的活性顯著增高。人類的端粒酶有三個亞基,包括人端粒酶RNA、端粒酶相關蛋白1和 hTERT,hTERT是端粒酶活性的決定因素。其表達水平在腫瘤中增加而在正常細胞中幾乎沒有表達[3]。
Horikawa等[5]克隆了hTERT上游調控區,發現該區域缺乏典型真核啟動子的核心元件TA TA盒和CAAT盒而富含GC,具有強啟動子活性,并與端粒酶的活性一致。hTERT啟動子中ATG上游-208~-150 bp的59個堿基是hTERT啟動子轉錄活性的基本單位,在轉錄起始位點上游含有幾個公認的細胞增殖和腫瘤發生相關的轉錄因子的結合位點,包括3個SP1元件,2個 E box及數個Myc相關鋅指蛋白(Myc-associated zinc finger protein,MAZ)和激活蛋白-2(activator protein-2,AP-2)結合位點[5]。先前的研究證實myc-max異二聚體就是與E box結合而激活hTERT啟動子的轉錄[10]。對啟動子CpG島的研究顯示其甲基化水平與啟動子活性有關[11]。以上研究說明hTERT的表達是在轉錄水平被調控的。
SUR是1997年由Ambrosini等[6]發現的一種細胞凋亡抑制因子,具有腫瘤特異性表達的特性。Bao等[12]證實SUR蛋白在腫瘤組織中的表達調控是發生在轉錄水平,因此認為,SUR基因的啟動子可能是一種腫瘤特異性的啟動子,能夠選擇性地在肝癌組織中激活其后的外源基因的轉錄,從而實現對治療基因在肝癌中的選擇性表達,避免損傷周圍正常肝組織。
Li等[13]通過分析Hela細胞中SUR基因的調控區,證實在SUR基因起始密碼子上游約0.4 kb的片段具有基本的啟動子活性。該區域缺乏TATA盒,但在基因讀碼框上游含有一段GC豐富的區域。在上游-6、-12、-171位,有 3個CDE元件(cell-cycle dependent element),在-42位,有 1個CHR區域(cell-cycle genes homology region)。有證據表明,上述元件在轉錄水平參與SU R在G2-M期特異性表達的調控。這與之前多項研究已經證實了的SUR的表達受細胞周期的調控,在G2-M期高表達相符。因此具有高度增殖活性的腫瘤細胞能夠比正常細胞表達更多的SUR轉錄產物[6]。
最近,Zhu等[14]研究證實,SU R啟動子在人類惡性黑素瘤、乳腺癌、卵巢癌的數個細胞系中具有相當高的啟動子活性,而在正常的人纖維母細胞和乳腺上皮細胞中其啟動子活性相當低,進一步證實了SUR啟動子具有腫瘤特異性轉錄的特性。
在本研究中,擴增了hTERT基因翻譯起始位點上游約0.3 kb的片段、SUR基因翻譯起始位點上游約0.4 kb的片段和AFP編碼區上游約0.3 kb的片段,結果證實:自人HepG2細胞克隆得到的啟動子序列與GenBank中源于其他腫瘤細胞的hTERT、SUR和AFP啟動子序列完全相同,說明這3種基因的基本啟動子在不同腫瘤細胞中具有高度保守性。
實驗證實在人肝癌細胞中AFP、hTERT和SUR啟動子均具有腫瘤特異性轉錄活性。將hTERT和SUR啟動子與AFP啟動子活性相比較,本研究發現,SUR和hTERT啟動子在肝癌細胞系中具有較AFP啟動子更強的轉錄活性,即使是在高表達AFP的HepG2細胞中,其啟動活性也比AFP啟動子活性高數十倍。結合SUR和hTERT在肝癌中極高的陽性率,本研究認為,利用SUR和hTERT啟動子調控治療基因轉錄,可望實現對肝癌靶向性的基因治療。
[1]Hallenbeck PL,Chang YN,Hay C,et al.A novel tumorspecific replication-restricted adenoviral vector for gene therapy of hepatocellular carcinoma[J].Hum Gene Ther,1999,10(10):1721.
[2]Huang X,Zhang W,Wakimoto H,et al.Adenovirus-mediated tissue-specific cytosine deaminase gene therapy for human hepatocellular carcinoma with different AFP expression levels[J].J Exp Ther Oncol,2002,2(2):100.
[3]Soder AI,Hoare SF,Muir S,et al.Amplification,increased dosage and in situ expression of the telomerase RNA gene in human cancer[J].Oncogene,1997,14(9):1013.
[4]Toshikuni N,Nouso K,Higashi T,et al.Expression of telomerase-associated protein1 and telomerase reverse transcriptase in hepatocellular carcinoma[J].Br J Cancer,2000,82(4):833.
[5]Horikawa I,Cable PL,Afshari C,et al.Cloning and characterization of the promoter region of human telomerase reverse transcriptase gene[J].Cancer Res,1999,59(4):826.
[6]Ambrosini G,Adida C,Altieri DC.A novel anti-apoptosis geng,survivin,expressed in cancer and lymphoma[J].Nat Med,1997,3(8):917.
[7]Ikeguchi M,Ueta T,Yamane Y,et al.Inducible nitricoxidesynthase and surviving messenger RNA expression in hepatocellular carcinoma[J].Clin Cancer Res,2002,8(10):3131.
[8]Moriai R,Asanuma K,Kobayashi D,et al.Quantitative analysis of the anti-apoptotic gene surviving expression in malignang haematopoietic cells[J].Anticancer Res,2001,21(1B):595.
[9]Ito T,Shiraki K,Sugimoto K,et al.Survivin promotes cell proligeration in huaman hepatocellular carcinoma[J].Hepatology,2000,31(5):1080.
[10]Wu KJ,Grandori C,Amacker M,et al.Direct activation of TERT transcription by c-MYC[J].Nat Genet,1999,21(2):220.
[11]Devereux TR,Horikawa I,Anna CH,et al.DNA methylation analysis of the promoter region of the human telomerase reverse transcriptase(hTERT)gene[J].Cancer Res,1999,59(24):6087.
[12]Bao R,Connolly DC,Murphy M,et al.Activation of cancer-specific gene expression by the survivin promoter[J].J Natl Cancer Inst,2002,94(7):522.
[13]Li F,Altieri DC.T ranscriptional analysis of human survivin gene expression[J].Biochem J,1999,344(Pt 2):305.
[14]Zhu ZB,Makhija SK,Lu B,et.al.Transcriptional targeting of tumors with a novel tumor-specific survivin promoter[J].Cancer Gene Ther,2004,11(4):256.
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
