重組凝乳酶搖瓶發酵條件的初步優化研究
2009-01-15 09:03:36張桂芝武彬黃保華劉雪蘭石天虹井慶川劉雪蘭魏祥法
山東農業科學 2009年12期
張桂芝 武 彬 黃保華 劉雪蘭 石天虹 井慶川 劉雪蘭 魏祥法
摘 要:凝乳酶是一種酸性蛋白酶,用作動物飼料添加劑,可提高動物特別是幼齡動物對飼料中蛋白成分的消化吸收率,促進其生長。本試驗研究了由畢赤酵母胞外表達多拷微小毛霉凝乳酶的搖瓶發酵條件。確定最佳表達條件為:甲醇每24 h添加1.0%,無氨基酵母氮源培養基(YNB)添加1.5%,采用pH 6.0的磷酸緩沖液,誘導120 h,在此條件下,重組凝乳酶的產量可達6 500 IU/ml。
關鍵詞:凝乳酶;發酵條件;畢赤酵母
中圖分類號:S816.79 文獻標識號:A 文章編號:1001-4942(2009)12-0081-04
凝乳酶(EC3.4.23.4)是一種酸性蛋白酶,能促進蛋白質凝結,用作動物飼料添加劑,可提高動物特別是幼齡動物對飼料中蛋白成分的消化吸收率,促進其生長[1]。幼齡動物的消化系統發育還不完善,對飼料中營養的消化吸收率較低。提高飼料中蛋白類的利用效率,不僅能夠節約飼料資源,降低養殖成本,而且能夠降低動物糞便的氨氮排放,減輕集約化飼養對環境的污染[2]。
巴斯德畢赤酵母(Pichia pastoris)表達系統是一種比較理想的真核微生物表達系統,是用于表達重組蛋白的標準工具之一,近年發展和應用較多[3]。Pichia pastoris具有與真核生物極其相似的分泌途徑和很強的真核蛋白質修飾功能,其自身分泌的蛋白質很少且易于高密度發酵,因此在表達和分離純化異源蛋白質等方面具有很強的優勢。但巴斯德畢赤酵母表達系統也會出現分泌蛋白聚合、降解等現象,從而降低蛋白的生物活性和產量[4]。此外,酵母細胞的生理狀態也會嚴重影響其產酶活性,培養基的pH和甲醇、無氨基酵母氮源培養基(YNB)等營養物質濃度,對細胞的代謝狀態會產生很大影響。本研究室成功地將微小毛霉凝乳酶在畢赤酵母中表達[5]。本試驗對重組凝乳酶發酵培養基的pH值、甲醇和YNB濃度,以及接種比例等發酵條件進行了優化,提高了重組凝乳酶的產量。
1 材料與方法
1.1 材料
菌種:具有12拷貝基因的整合重組畢赤酵母菌PIC9K-mcp-03。
培養基:BMGY/BMMY培養基:取酵母提取物10 g,蛋白胨20 g,溶于700 ml去離子水中,高壓滅菌20 min,冷卻至室溫,加入1 mol/L磷酸鉀緩沖液(pH 6.0)100 ml、10×YNB 100 ml、500×B 2 ml、10×GY 100 ml(若制備BMMY,用100 ml 10×M代替10×GY),4℃保存備用(如無特殊說明,均采用上述條件,改變其中某項發酵條件時,其它項目不變)。1 000 ml三角瓶中加入100 ml培養基,于29℃ 170 r/min振蕩培養5 d,每24 h補加甲醇1次。
1.2 方法
1.2.1 甲醇和YNB添加量對發酵液的影響 改變BMMY培養基中甲醇(0~3%)和YNB(0~4%)的添加量,其它條件不變,培養120 h,測定發酵液OD600 nm和發酵上清液酶活力。
1.2.2 接種量對重組凝乳酶產量的影響 改變BMMY培養基的接種量,選擇不同體積的BMGY培養獲得的菌體細胞,加入BMMY培養基中培養120 h,測定發酵液OD600 nm和發酵上清液酶活力。
1.2.3 BMMY培養基的初始pH值對重組凝乳酶產量的影響 改變BMMY培養基中所含磷酸緩沖液的pH值,分別調至pH 4.0~7.0,培養120 h,測定發酵液OD600 nm和發酵上清液酶活力。
1.2.4 菌體濃度的計算方法 將發酵液離心,菌體沉淀以蒸餾水重懸,經適當稀釋后,測定并計算菌體懸濁液在600 nm的OD值。
1.2.5 酶活測定采用Arima方法[7] 適當稀釋的上清液0.5 ml 35℃保溫5 min,加入到35℃預保溫10 min的5 ml 10%脫脂牛乳中,計時至管壁出現顆粒。記錄凝乳時間(T)。酶活定義:在一定溫度下(35℃),40 min凝乳1 ml牛乳的酶量為一個酶活單位。
凝乳酶活力(U)=[(5×2400)/(0.5×T)]×n,其中:T為凝乳時間,n為稀釋倍數。
2 結果與分析
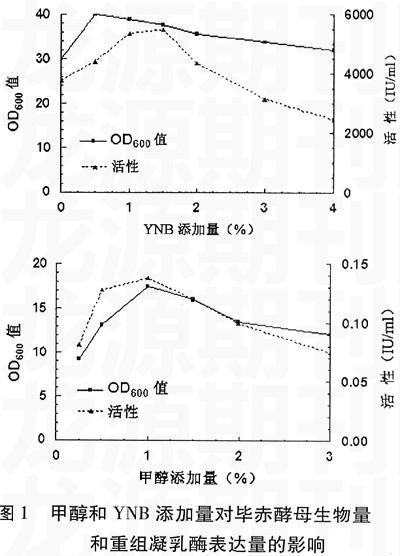
2.1 甲醇和YNB添加量對重組凝乳酶產量的影響
甲醇和YNB在培養基中起提供營養的作用,甲醇除了是發酵培養基的碳源,還是外源蛋白表達的誘導物,其濃度對重組凝乳酶的表達有一定的影響,隨著甲醇濃度的增加重組凝乳酶的表達量也增加。而YNB除了作為培養基的氮源,還含有維生素和多種無機鹽類。但二者在較高濃度下,又對發酵有抑制作用,且高濃度的甲醇對酵母菌也有毒害作用,甲醇經代謝生成甲醛和甲酸,甲醛可與帶有羥基、巰基、氨基基團的分子發生親核加成反應,向其提供亞甲基,使自由的分子鏈被甲醛交聯起來,失去原有的生理功能,因此會影響重組凝乳酶的表達量。而YNB中含有的(NH4)2SO4、K3PO4、和NaCl、CaCl2等無機鹽在較高濃度下,同樣會影響畢赤酵母的生長代謝,降低重組凝乳酶的表達量。培養基中YNB和甲醇的添加量對重組凝乳酶的影響如圖1所示。結果表明甲醇濃度在1%、YNB濃度為1.5%時,畢赤酵母的生物量與重組凝乳酶的產量達到最高值。
2.2 接種量對重組凝乳酶產量的影響畢赤酵母的接種量實際上是有可能表達重組
凝乳酶的細胞數,這些酵母是否表達目的蛋白,還受多種環境因素的影響。除了底物濃度之外,實際上培養基中的溶氧量具有重要的影響。圖2為接種量影響重組凝乳酶表達量的結果。隨接種量的增加,菌體生物量與酶活力的變化情況并不一致,酶活力在接種量為20%時達到最高值,25%也保持較高水平,之后略有下降;而菌體生物量在接種量為30%時達到最高,之后迅速下降。因此,溶氧量很可能是畢赤酵母表達的主要限制性因素。因為溶氧量受多種因素影響,改變搖床的轉速、培養基成分等都會改變溶氧濃度,并導致表達量的改變。因此,最佳接種量的測定會隨上述條件改變而略有波動,一般在16.7%~25.0%之間。

2.3 培養基的初始pH值對重組凝乳酶產量的影響
改變培養基的pH值對細胞的活性也會產生很大影響。重組凝乳酶在pH 3.2~8.0都比較穩定,因此發酵培養基可選的pH值范圍較廣。根據酵母的代謝特點,測定了初始培養基在pH 4.0~7.0畢赤酵母的生物量和凝乳酶的表達量(圖3)。在發酵過程中酵母會產酸,因此改變了加入培養基的磷酸緩沖液的pH值。

酵母表達系統中外源蛋白的降解失活和不同pH值對酵母生理狀態的影響,是導致重組凝乳酶表達量的兩項主要因素。圖3的結果顯示,在pH 4.0~7.0范圍內,凝乳酶的活力與菌體濃度變化基本一致,推測可能是由于重組凝乳酶在不同pH值下穩定性較強,且凝乳酶自身作為蛋白酶具有較強的蛋白酶水解抗性。
3 討論與結論
發酵時間、營養和誘導物的添加量、接種量以及培養基初始pH值對重組凝乳酶的產量都有顯著影響,但上述影響只是表觀現象。實際上,這些因素的影響更深入地體現在對酵母的生理狀態和重組凝乳酶的特定影響兩個方面。
甲醇作為發酵的誘導物和碳源,而YNB作為氮源,在一定濃度下對重組凝乳酶的表達具有促進作用,但二者在較高濃度下,又對發酵有抑制作用。實際上不論是甲醇還是YNB對畢赤酵母生物量和表達量的影響并不一致。在低濃度下,甲醇主要表現誘導作用,而只表現出較弱的作為碳源的作用。因此,甲醇添加量為0.5%時,盡管OD600nm表征的菌體量較低,卻獲得了較高的酶活力。而在添加量為0.5%時,YNB作為氮源則主要表現出對菌體的促進生長作用,只有在滿足了菌體生長需要后,在添加量為1.0%~1.5%時,才表現出促進重組蛋白分泌的作用。
接種量以及營養物添加濃度導致重組凝乳酶產量的變化,從根本上講,體現的都是酵母生理狀態對重組蛋白分泌量的影響。OD600 nm所表征的菌體濃度與蛋白分泌量的差別正說明了這一點。以甲醇為碳源,需要消耗大量的溶解氧,提高接種量,菌體濃度增加,勢必降低單位細胞可利用的溶解氧含量,從而降低單位細胞重組蛋白的表達量。
酸堿度對蛋白表達量的影響主要體現在兩個方面:首先,發酵液初始pH值將影響酵母細胞的生理狀態;其次,發酵液的pH值還對表達蛋白的穩定性有一定的影響。因為凝乳酶具有較強的pH值穩定性,而且該酶本身為真菌胞外蛋白酶,對其它蛋白酶的水解具有一定的抗性,所以,圖3顯示的培養基初始pH值對畢赤酵母生物量和表達量的影響比較一致,即本試驗中,發酵液初始pH值影響重組凝乳酶表達量,主要是源于對酵母細胞生理狀態的影響。
上述各種因素對酵母細胞的生理狀態具有不同的影響機理,各種影響作用相互交叉、十分復雜,而正交試驗設計要求各個影響因素的作用彼此相互獨立。因此,本試驗的各項影響因素不宜簡單套用正交試驗進行最佳發酵條件的優化,而試驗中獲得的發酵條件還有進一步優化的空間。
參 考 文 獻:
[1] 肖 競,孫建議,李衛芬.酸性蛋白酶及其在畜牧業中的應用[J].飼料博覽, 2003, 3:27-29.
[2] 費笛波,錢玉英,葉玲玲,等. 黑曲霉菌株6042飼用酸性蛋白酶的性質及其應用研究[J].浙江農業學報,1997, 9(6):300-304.
[3] Hao Y Y, Chu J,Wang Y H,et a1.Expression and aggregation of recombinatant human consensus interferon-α mutant by Pichia pastoris[J]. Biotechnology Letters, 2006, 28(12):905-909.
[4] Macauley-Patrick S,Fazenda M L,McNeil B,et a1.Heterologous protein production using the Pichia pastoris expression system[J].Yeast, 2005, 22: 249-270.
[5] 張桂芝,武 彬,井慶川, 等.微小毛酶凝乳酶的基因克隆及在畢赤酵母GS115中的表達[J].山東農業科學,2009,11:1-4.
