木霉菌REMI轉化體對番茄灰霉病的防治及其機理的研究
2008-04-29 00:44:03劉限郭培磊高增貴趙巖
植物保護 2008年5期
關鍵詞:利用
劉 限 郭培磊 高增貴 趙 巖
摘要:研究不同木霉菌轉化體對番茄灰霉病防治效果及機理,為木霉菌生物防治的合理利用奠定基礎。利用限制性內切酶介導基因整合技術(restriction enzyme mediated integration,REMI),通過插入線性化質粒DNA獲得了生物防治番茄灰霉病(Botrytis cinerea)效果優于出發菌T21菌株(出發菌)的3個木霉菌轉化體Ttrm31、Ttrm34和Ttrm55,對侵染花器和葉片的灰霉病防效分別比原生物防治木霉菌株提高了16.9%和8%。木霉菌轉化體的產孢能力、分生孢子的萌發率、對碳氮源的利用能力及對高溫的抵抗能力都有所提高;木霉菌轉化體本身產生的幾丁質酶和β-1,3-葡聚糖酶的活性均比出發菌高,因此通過REMI技術可以獲得新的有益木霉菌轉化體,在一定程度上提高了生物菌株防治番茄灰霉病的水平。說明REMI技術可以用于改良生防木霉菌株的功能,提高生物防治效果。
關鍵詞:限制性內切酶介導整合技術;木霉菌轉化體;番茄灰霉病;生防機理
中圖分類號:S 482.2.92,S 436.412.13
保護地的高濕、適溫條件為番茄灰霉病(Botry-tis cinerea Pers.ex Tris.)的發生創造了適宜的環境,因而該病害成為國內外工廠化高效農業番茄生產中的主要障礙。由于目前生產上仍缺少抗病品種,多數農藝性狀優良的品種對灰霉病高度感病,且灰霉病菌已對多種化學殺菌劑(多菌靈等)產生了明顯的抗藥性,嚴重制約了番茄的安全生產。因此構建對保護地蔬菜各種逆境均具有明顯抗性,同時又能提高對番茄灰霉病抗性的工程菌株具有重要意義,也是今后木霉菌能否成為蔬菜葉部病害無公害防治有效手段的關鍵。本文以生物防治番茄灰霉病菌的木霉菌(Trichoderma sp.)T21菌株為對象,探索利用REMI技術改良的生物防治菌株對番茄灰霉病的防治效果及可能的生防機理,為我國生物農藥工程菌株創建提供新技術,為番茄灰霉病的生物防治提供高效菌株。
1材料與方法
1.1材料
番茄品種為L402。
供試木霉菌為野生菌株T21和轉化體Ttrm31、Ttrm34、Ttrm55。灰霉病菌由沈陽農業大學植物免疫室生物工程中心保存。
1.2方法
1.2.1活體植株防病效果測定
選擇發育齊一的番茄植株,接種前澆足水。選擇3株離體條件下抑病效果較好的木霉菌轉化體(Ttrm31、Ttrm34和Ttrm55),采用孢子懸浮液噴霧的方法接種。每處理10株,30株噴孢子懸浮液150 mL,然后采用隨機擺放的方法將3個處理擺放于人工氣候室中,用補濕器補濕,將濕度保持在90%以上,24 h后接種灰霉病菌,7 d后調查發病情況。
1.2.2木霉菌轉化體的孢子萌發
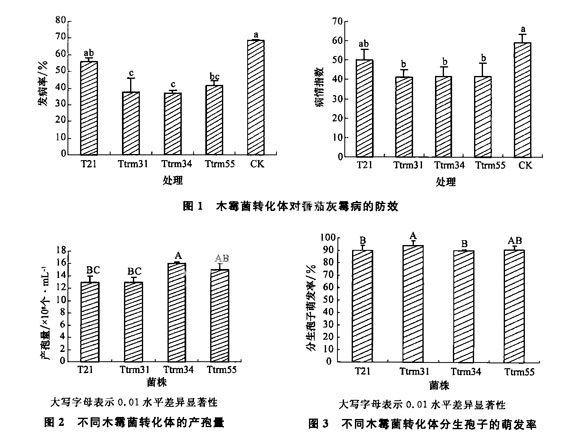
將5%葡萄糖的瓊脂平鋪在無菌的載玻片上,凝固后將調整到一定濃度(103個/mL)的孢子懸浮液滴加到瓊脂上,在無菌的條件下吹干后,將載玻片放在含有水分的濾紙上,于培養皿中25℃下培養12~15 h后,在顯微鏡下調查孢子的萌發情況。
1.2.3木霉菌轉化體的產孢能力
將含有木霉菌孢子的菌片轉移到PDA培養基的中央,在28℃下培養7 d后,用10 mL無菌水將孢子洗至試管中,充分振蕩使孢子分散均勻,然后用血球計數板計數孢子的數量,從而計算孢子懸浮液的濃度,確定不同轉化體的產孢能力。
1.2.4木霉菌轉化體對碳氮源的利用能力
以葡萄糖20 g,天冬氨酸2 g,KH2PO41 g,Mg-SO4·4H2O 0.05 g,FeSO4·7H2O 0.01 g,MnSO4·4H2O 3.2 mg,ZnSO4·7H2O 1.8 mg,瓊脂20 g,水1 000 mk,分別用等量的蛋白胨、硝酸鈉和尿素代替天冬氨酸,制成不同氮源的培養基。將轉化體的菌片轉移到培養皿的中央,于28℃下培養,48 h測量菌落半徑。
以葡萄糖20 g,天冬氨酸2 g,KH2PO41 g,Mg-SO4·4H2O 0.05 g,FeSO4·7H2O 0.01 g,MnSO4·4H2O 3.2 mg,ZnSO4·7H20 1.8 mg,瓊脂20 g,水1 000 mL,分別用等量的蔗糖、淀粉、麥芽糖代替葡萄糖,制成不同碳源的培養基。將轉化體的菌片轉移到培養皿的中央,于28℃下培養,每隔48 h測量菌落半徑。
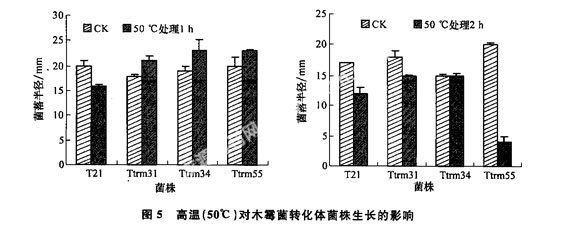
1.2.5高溫對轉化體生長的影響
將培養3~4 d的轉化體和T21木霉菌菌片放置到PDA培養基上,在50℃條件下放置1~2 h,然后再轉移到正常溫度(25℃)下培養,30~40 h后測量菌落半徑。
1.2.6木霉菌轉化體幾丁質酶和β-1,3-葡聚糖酶活性的測定
幾丁質酶的測定參照顧向陽、陳捷等方法,略加改進。β-1,3-葡聚糖酶活測定參照杜良成的方法,以葡聚糖為底物,根據從葡聚糖中釋放出葡萄糖的量來測定酶活性。
2結果與分析
2.1木霉菌轉化體對番茄灰霉病防治效果
采用限制性內切酶介導整合(REMI)技術,獲得木霉菌轉化體并應用于番茄灰霉病防治,通過對峙培養和離體葉片試驗篩選出3株對灰霉病菌生防效果好的木霉菌轉化體菌株Ttrm31、Ttrm34和Ttrm55。然后將此3株菌株在整株番茄上進行了對番茄灰霉病的防治試驗。結果表明,所有番茄均已發病,發病率都達到了35%以上。轉化體菌株Ttrm31、TtRm34和Ttrm55的發病率明顯低于野生菌株T21和對照。發病率最低的為轉化體菌株Ttrm34,發病率為36.9%。所有木霉菌轉化體對番茄灰霉病的防效比野生菌株T21的防效都有所提高,其中轉化體菌株Ttrm31對番茄灰霉病的防效是最好的,其病情指數為41.1,但是該菌株和菌株Ttrm34和Ttrm55的防效差異不明顯,菌株Ttrm34和Ttrm55的病情指數均為42.5;而野生菌株T21的病情指數為50.4;對照的病情指數為593。從中可以看出,木霉菌轉化體對番茄灰霉病的防效有了明顯的提高(圖1)。
2.2木霉菌轉化體生防活性機理的研究
2.2.1不同木霉菌轉化體的產孢情況
由圖2看出,不同轉化體產孢能力的差異達到
了極顯著水平。Ttrm34產孢能力最高,極顯著高于野生菌株T21,而轉化體Ttrm31和Ttrm55的產孢能力基本上與野生菌株T21一致,都達到了109個/mL。說明REMI技術插入的位點不同,從而影響了轉化體的產孢能力。
2.2.2木霉菌轉化體分生孢子的萌發
由圖3看出,不同轉化體的分生孢子萌發率有極顯著(P=0.01)的差異,相對于野生菌株T21,轉化體的孢子萌發率發生了很大變化,其中Ttrm31分生孢子的萌發率最高,達到了94%,顯著高于T21;而Ttrm55和Ttrm34的分生孢子的萌發率分別為91%和90%,與野生菌株T21(90%)基本一致。
2.2.3木霉菌轉化體對碳氮源的利用能力
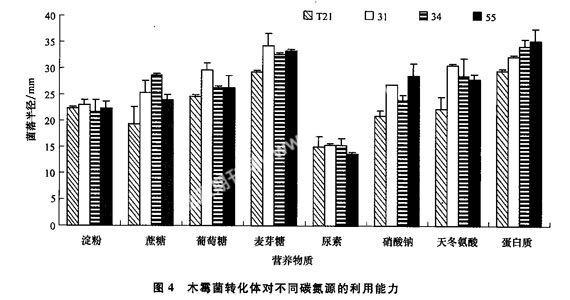
由圖4可以看出,木霉菌轉化體對不同氮源的利用能力不同。Ttrm31、Ttrm34和Ttrm55可以很好地利用各種氮源,在尿素為氮源的培養基上12 h時就可以生長,而野生菌株T21菌株則不能生長;在硝酸鈉、蛋白質和天冬氨酸為氮源的培養基上Ttrm31、Ttrm34和Ttrm55的生長速度大于野生菌株T21。另外木霉菌利用有機氮源的能力較強,而對無機氮源的利用能力較弱。總體來說,轉化體對各種氮源的利用能力好于野生菌株T21,說明RE-MI插入引起了轉化體對氮源利用的差異。不同木霉菌菌株對碳源的利用能力也不同,在各種糖類為碳源的培養基上,3株木霉菌轉化體的生長速度都快于野生菌株T21,說明Ttrm31、Ttrm34和Ttrm55能夠充分利用各種碳源,生長速度快,說明REMI插入引起了轉化體對碳源的利用能力不同。另外木霉菌對不同碳源的利用能力也不同,能夠很好地利用麥芽糖、葡萄糖和蔗糖,而對淀粉的利用能力較弱。

另外從菌絲的稀疏度也可以看出不同菌株對各種營養的利用存在差異(表1)。不同菌株在相同的培養基上菌絲的稀疏度存在差異,如,在葡萄糖作為碳源的情況下,Ttrm55的菌絲比較稀疏,而其他木霉菌轉化體和野生菌株的菌絲則正常;在以天冬氨酸為氮源的情況下,個trm55的菌絲比較密,而其他木霉菌轉化體和野生菌株的菌絲則相對較稀疏。在其他氮源和碳源的培養基上生長的菌絲也存在菌株間稀疏度不一樣的現象。說明不同菌株對各種養分利用能力不同,也間接說明了轉化體是由于質粒插入的位點不同造成的,說明了有外源的基因插入。
從轉化體在各種不同氮源和碳源培養基上培養性狀可以看出,不同轉化體對營養物質的利用能力及菌落形態和顏色均不同。
2.2.4木霉菌轉化體對高溫的耐性變化
試驗表明,所有轉化體和野生菌株T21在50℃下處理1~2 h后再正常培養,均能保持正常生長,而且50℃下處理1 h,還能促進轉化體生長,但在該溫度下處理2 h以上,轉化體和野生菌株T21的生長、產孢及菌落形態將受到一定的影響。由于45~50℃下處理時間不超過1 h可保證菌株正常生長,因此在夏天或者春天悶棚中應用轉化體時不會影響生防活性的發揮(圖5)。總之在高溫處理不超過2 h的情況下,轉化體和野生菌株可以生長,而且轉化體在高溫處理1 h時,生長速度反而加快了。
2.2.5酶活性的變化
2.2.5.1幾丁質酶活性
由圖6可以看出,不同轉化體產生的幾丁質酶的活性不同,其中Ttrm34的活性最高,其次為Ttrm55,野生菌株T21的活性最低。轉化體的幾丁質酶活性普遍升高,說明由于REMI插入的存在,激活轉化體幾丁質酶基因,使幾丁質酶表達量增加。但是通過F檢測,沒有達到顯著水平上的差異。
2.2.5.2β-1,3-葡聚糖酶活性
由圖7可以看出,不同轉化體產生的β-1,3-葡聚糖酶的比活性不同,其中Ttrm34的活性最高,其次為Ttrm55,野生菌株T21的活性最低。轉化體的β-1,3-葡聚糖酶活性提高,說明轉化體中插入了質粒pV2,造成β-1,3-葡聚糖酶基因的激活,但是通過F檢測,沒有達到顯著水平上的差異。同時發現該酶與幾丁質酶具有一定相關性。
3討論
3.1REMI技術可作為創造木霉菌變異獲得生物防治新菌株的手段之一
由于限制性內切酶消化片段對染色體的插入是隨機的,即可能發生在無意義序列、調控序列或閱讀框架內的一個或幾個識別位點上,質粒依賴其相容末端插入被切開的基因組中。一旦整合于有意義序列,該序列控制的一系列性狀將產生突變。其插入的隨機性決定著受影響的表型將是多種多樣的。目前REMI技術主要應用于不同植物病原菌致病相關基因的研究中,且有用于創造木霉菌株變異、篩選新生防菌株的報道。本文在優化木霉菌T21菌株原生質制備、再生、轉化條件的基礎上,利用REMI技術將外源質粒DNA隨機插入到木霉菌的染色體中,從而使木霉菌中某些基因激活或者失活,篩選出幾株生防機理不同的木霉菌轉化體。
3.2REMI技術可改變生物防治菌株的多種機能
研究中發現REMI技術可改變出發菌T21原來的一些生物學特性,如Ttrm34產孢量極顯著增加;Ttrm31分生孢子萌發率極顯著提高;各突變株利用碳氮源的能力有所提高;菌株對高溫的抗性也發生一定的變化,50℃高溫處理1 h可促進木霉菌轉化子的生長,而出發菌T21在此溫度下生長受到了一定的抑制。這些變化對生防活性的提高都有一定的貢獻,由此看出木霉菌生防活性不是哪一種機理能達到的,需要從多方面來研究。總之,REMI技術如結合適宜的木霉菌轉化體篩選技術,有希望成為獲得多種生物防治功能菌株或發現新生物防治機理的方法。
3.3木霉菌優良轉化子為生物防治番茄灰霉病提供了新途徑
本文從REMI技術構建的木霉菌轉化子中篩選出防效明顯提高的生防菌株,與出發菌相比轉化子對番茄花器和葉片灰霉病的防效平均提高了16.9%和8.0%。由于在溫室內灰霉病對花器侵染后會進一步侵染果實,從而影響番茄的產量,因此對花器侵染的控制,必將降低果實灰霉病的發生,對提高田間防效更有意義。篩選的轉化子多代繁殖后仍具有穩定的抗潮霉素性能,說明REMI技術構建的轉化子在一定條件下能實現不可逆遺傳改良。這反映出外源基因片段誘發木霉菌發生有益變異的潛力較大,為今后進一步利用木霉菌插入變異潛力,開發多功能、高效生防因子奠定了新的技術基礎。當然在選擇過程中需要注意從多方面來綜合考慮木霉菌對番茄灰霉病的防治效果,不能只考慮一個方面。另外,根據木霉菌轉化體生防機理的不同,可以研究將多種木霉菌轉化體混合施用來提高木霉菌對番茄灰霉病的防治效果或許能夠改變木霉菌生防制劑活性低和不穩定的問題。
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
中學生數理化(高中版.高考數學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業設計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
