小麥抗條銹新品系89144抗銹機理研究
2006-04-29 04:11:09倪建福歐巧明令利軍葉春雷王亞馥邢更妹李杉崔凱榮武祿光
植物保護 2006年2期
倪建福 歐巧明 令利軍 葉春雷 王亞馥 邢更妹 李 杉 崔凱榮 武祿光
摘要將高粱總DNA通過花粉管通道導入小麥感病品種甘麥8號,D2代出現2株對條銹病免疫的變異株,D5代有9個株系抗性已經穩定;用混合菌和分小種鑒定,對條中29、30、洛13Ⅱ、水14、水14中梁17-s、HY3、條中31號等小種表現免疫。結果分析表明,新品系89144接種后組織中SOD活性升高,原受體甘麥8號接種銹菌后SA含量也有升高;但并不伴隨有CAT活性下降,SOD活性和H2O2含量的升高;推測5A為CAT過氧化活性提供一個電子的過程中SA含量必須達到一定的閾值,并且與CAT的時序調節相配合;據SA結合態和游離態含量的變化動態,表明89144具有SA信號傳導途徑,推測SA信號傳導途徑的上游應該還有一些機制在起作用。
關鍵詞植物病理學:小麥條銹病;抗銹新品系
中圖分類號S435.121.42
小麥條銹病(Puccinia striiformis f.sp.triti-ci)是甘肅省小麥主要病害之一,嚴重影響小麥高產穩產。國內外研究和生產實踐證明,選育抗銹病品種是防治該病最經濟有效的途徑。為了培育新的抗銹品種,從1988年起,在總結以往工作的基礎上開展了應用花粉管通道法將外源C4作物高粱DNA向小麥轉移的探索,1989年在大田和溫室共做了80個外源DNA導人組合,獲得抗條銹病新品系89144,并對其后代材料作了較系統的觀察和研究。本文報道小麥抗條銹新品系主要特征和抗性生理生化變化進行比較測定和分析。
1材料和方法
1.1材料
受體為普通小麥(Triticum sestrvum),品種甘麥8號,株高107cm左右,穗型棍棒狀、頂芒、生育期104d,適應性強,曾大面積種植,現重感條銹病;供體為高粱[Sorghumbicolor(L.)]品種米高粱,抗旱、耐澇、耐鹽堿、適應性強。
1.2方法
1.2.1DNA提取和導入
供體高梁DNA提取和導人參照倪建福等方法。
1.2.2田間資料統計與觀察
經處理的當代受體記作DO,其后代依次記為D1、D2和D3等。DO~D1按組合收種子,以后按系譜法選擇,等抗性穩定后再按組合收種子,種植按行長1.20m、行距0.20m、粒距0.04 m點播,田間觀察和室內外考種項目按常規育種要求進行,各種材料按種植小區和株系收獲。
1.2.3抗銹性鑒定
除本中心每年用心葉涂抹法接種流行混合菌選擇外,對獲得的抗銹新品系再送甘肅省農業科學院植保所進行專門鑒定,反應型按常規標準匯載。
1.2.4種子粗蛋白和淀粉含量的測定
抗銹新品系89144和供、受體一并送測試中心,按照國家規定的標準方法測定。粗蛋門是將材料先采用凱氏法處理后,用瑞典Tecalor公司的Kielteasystem Ⅱ氮/蛋白質分析儀測定;賴氨酸采用國際染料結合法(PBL法)染色,再用北京環保儀器廠的GXDL。-202型蛋白質賴氨酸分析儀測定;淀粉采用國際旋光法測定”。
1.2.5水楊酸(psalicylicacid,SA)含量的測定
將小麥種子浸種后,播種于塑料缽蛭石內,在小麥一葉展葉時利用心葉涂抹法接人小麥條銹菌混合菌種,培養溫度13~19℃.光照12h/d,濕度100%,接種后24、32、40、48、56 h取材。參考李兆亮等方法用硅膠板純化粗提液,用HPLC(儀器型號WatersTM600EC-18柱)檢測樣品中SA含量,檢測溫度40℃,流速為0.6 mL/min,檢測波長310 nm,保留時間4.05 min。
1.2.6過氧化氫酶(CAT)活性的測定
取0.5g葉片加1.8mL預冷的提取緩沖液(10 mmoI/L Tris-HCl pH7.5,0.25 mol/L蔗糖,1 mmol/L EDTA,10.5 mmol/L DTT,0.1 mmol/LPMSF),研磨成勻漿后在4℃條件下27000g離心15min,上清液按崔凱榮方法測定酶活性。
1.2.7過氧化氫(H2O2)含量的測定
取1g葉片參考Patterson等方法加3mL冷丙酮磨成勻漿,16000g離心10 min,上清液定容,反應液中含0.1mL體積分數20%TiCL4的濃鹽酸,0.2 mL濃氨水和1mL。上清液。生成的過氧化物-Ti復合物用丙酮洗5次.丙酮揮發后溶于3mL。硫酸(1mol/L)中,測410nm光吸收值,制作H2O2標準曲線。
1.2.8超氧化物岐化酶(SOD)活性的測定
取0.5g葉片加1,8 mI。預冷的提取緩沖液(50mmol/LpH7.8磷酸緩沖液,0.1 mmol/LED-TA,體積質量0.3%TritonX-100;體積質量4%聚乙烯吡咯烷酮(PVPP)研磨后用紗布過濾,10500g離心20 min,上清液為粗酶液。按照EL-moshaty等方法測定SOD活性,根據SOD抑制NBT光化學還原的量計算酶活性。
1.2.9過氧化物酶(POD)活性的測定
取0.5g葉片參考崔凱榮等方法加1.8 mL預冷的提取緩沖液(100mmol/LPBS,pH6.0.內含體積分數0.1%TritonX-100和體積質量0.04%偏重亞硫酸鈉。冰浴研磨,14 000 S離心15 mln。以愈創木酚作底物,在470 nm處測OD值。以△OD/min·mg蛋山為酶活單位。
2結果與分析
2.1生物學特性
1989年將高梁DNA通過花粉管通道導入小麥感病品種麥8號,共處理小花或子房134個,獲得Do代種子28粒,結實率為20.90%。1990年點播成苗27株;1991年種植900株,從中選出2株變異株;1992年從中選1株種子點播;1993年種植l800株.獲得變異株121株;1994年種植1140株,入選41株;1995年種植1800株,從中選出穩定遺傳的抗銹新品系89144系列,其生物學特性,見表1。
兩個抗銹新品系的大多數生物學性狀是介于受體和供休之間,如葉功能期延長,株高降低,穗長和粗蛋白增加以及千粒重、賴氨酸和淀粉含量等均介于供體和受體之間,但籽粒飽滿度增加,小穗排列疏松,葉片變窄變長,穎殼變硬(表2)。
抗銹性鑒定結果表明,兩個抗銹新品系89144
對目前流行的所有小種免疫,特別是在近年甘肅生產上應用的所有小麥品系幾乎對條中3l和32號喪失抗性的情況下,89144仍然抗銹,與其雜交的組合后代也非常抗銹,見表2。
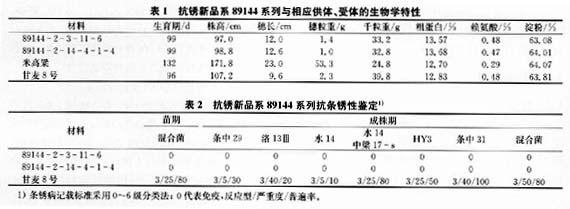
2.2水楊酸含量變化
在接種條銹菌后,通過HPLC測定結合態與游離態SA含量變化結果如圖l、2。抗銹新品系89144游離態水楊酸從32 h開始逐漸升高,40 h達到峰值,然后丌紿緩慢下降,到48h急劇下降。甘麥8號從24h開始緩慢上升,上升幅度比89144小2倍,到32 h快速下降,而在未接種銹菌的對照中,89144和甘麥8號內源游離SA含量變化都不大。圖2表明,接種后89144、甘麥8號的結合態SA含量都有上升,并且甘麥8號比89144上升幅度大而且時間早。在對照中,89144、甘麥8號的結合態SA含量變化不大。
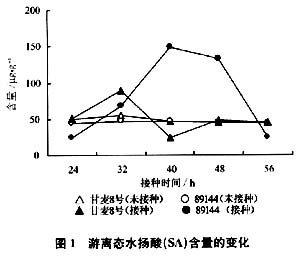
2.3過氧化氫酶活性變化
由圖3可以看出,抗銹新品系89144的CAT活性在接種條銹菌40h處開始大幅度下降,48h后又略有回升,而甘麥8號變化不大。對照未接種條銹菌的89144、甘麥8號,CAT活性變化不大。
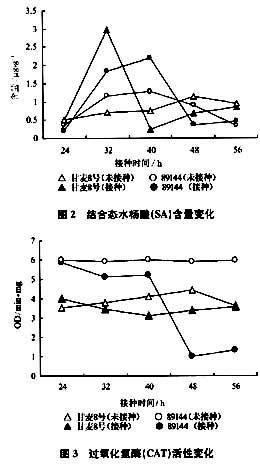
2.4過氧化氫含量變化
圖4表明,在接種銹菌48 h后,抗銹新品系89144中H2O2含量達到峰值,甘麥8號含量也略有升高,而對照89144、甘麥8號H2O2含量變化不大。
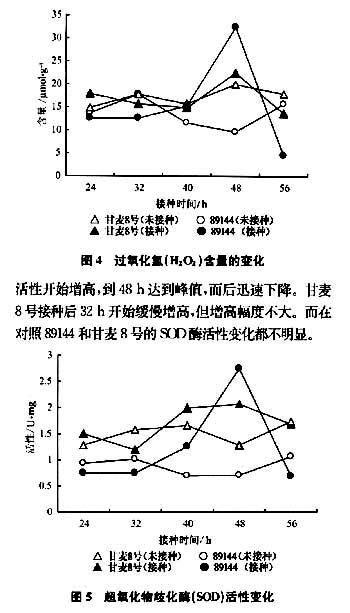
2.5超氧化物歧化酶活性變化
圖5表明,抗銹新品系89144在接種后40h,SOD活性開始增高,到48h達到峰值,而后迅速下降。甘麥8號接種后32h開始緩慢增高,但增高幅度不大。而在對照89144和甘麥8號的SOD酶活性變化都不明顯。
2.6過氧化物酶活性變化
圖6中抗銹新品系POD酶活性無論在接種條件下還是在未接種條件下變化都不大,只是在接種56 h后抗銹新品系89144的POD活性稍有升高。

3討論
經多年實踐表明,利用花粉管通道將外源DNA導入小麥胚囊,轉化上不具備正常細胞壁即合子或早期細胞是完全可行的,因為DNA片斷與受體基因組部分基因間有可能存在同源性而發生雜交重組。該技術之所以提高選育出抗銹品系的成功率,是因為供體一方的染色體經提取后片斷變小,從而減少了整條染色體之間的排斥性,增加了部分染色體重組幾率。
實驗研究顯示,外源DNA導入后代抗銹新品系89144在接種銹菌40h后,內源游離SA含量達到峰值,從40 h至48 h有一個緩慢下降階段,同時伴隨著CAT活性降低,H2O2含量增加,SOD活性增高。而不抗銹品種甘麥8號在接種銹菌后,也有內源游離SA含量升高,但并不伴隨有CAT活性下降和SOD、POD活性升高,也沒有H2O2含量的上升。Chen等曾研究證明煙草中的SA結合蛋白(SA—bindingprotem,SABP)具有CAT活性,可以降解H2O2產生O2和H2O,SA阻遏SABP的CAT活性,SA作為電子供體為CAT的過氧化活性提供一個電子,同時抑制并使這些酶處于不活躍狀態,從而導致H2O2含量升高。從本實驗結果可以看出雖然抗銹新品系89144和甘麥8號接種銹菌后,SA含量都有升高,但甘麥8號H2O2含量達到峰值的時間早,峰值低,這可能是因為SA為CAT過氧化活性提供一個電子的過程中SA的量必須達到一定的閾值,并且與CAT的時序調節相配合。
實驗中還發現,甘麥8號、抗銹新品系89144接種銹菌后游離SA含量增高,結合態SA含量也升高,所以在誘導條件下游離SA含量的升高不一定來自于結合態SA的釋放,還可能是植物體內與SA合成的相關基因被啟動。這樣,在SA信號傳導途徑的上游就還有某些機制在起作用。抗銹新品系89144接種銹菌后SOD活性也有升高,SOD可使O2通過歧化反應產生H2O2,在正常條件下,植物體中的SOD活性足以清除體內的O2,使O2保持在正常生理水平。但在逆境條件下,O2水平的提高刺激了SOD活性的增加,這可能與病原物誘導的氧化噴發有關。但在實驗中并沒有發現POD活性的大幅度變化,也許在檢測時POD活性已恢復常態,這也符合氧化噴發的時相。由此推測在植物體內H2O2含量升高可以通過多種途徑造成,Chen等的實驗也證明H2O2或它的衍生物活性氧可激活抗病反應途徑中相關基因表達。
